

Intracellular trafficking and endocytosis of CXCR4 in fetal mesenchymal stem/stromal cells
- PMID: 24885150
- PMCID: PMC4065074
- DOI: 10.1186/1471-2121-15-15
Intracellular trafficking and endocytosis of CXCR4 in fetal mesenchymal stem/stromal cells
Abstract
Background: Fetal mesenchymal stem/stromal cells (MSC) represent a developmentally-advantageous cell type with translational potential.To enhance adult MSC migration, studies have focussed on the role of the chemokine receptor CXCR4 and its ligand SDF-1 (CXCL12), but more recent work implicates an intricate system of CXCR4 receptor dimerization, intracellular localization, multiple ligands, splice variants and nuclear accumulation. We investigated the intracellular localization of CXCR4 in fetal bone marrow-derived MSC and role of intracellular trafficking in CXCR4 surface expression and function.
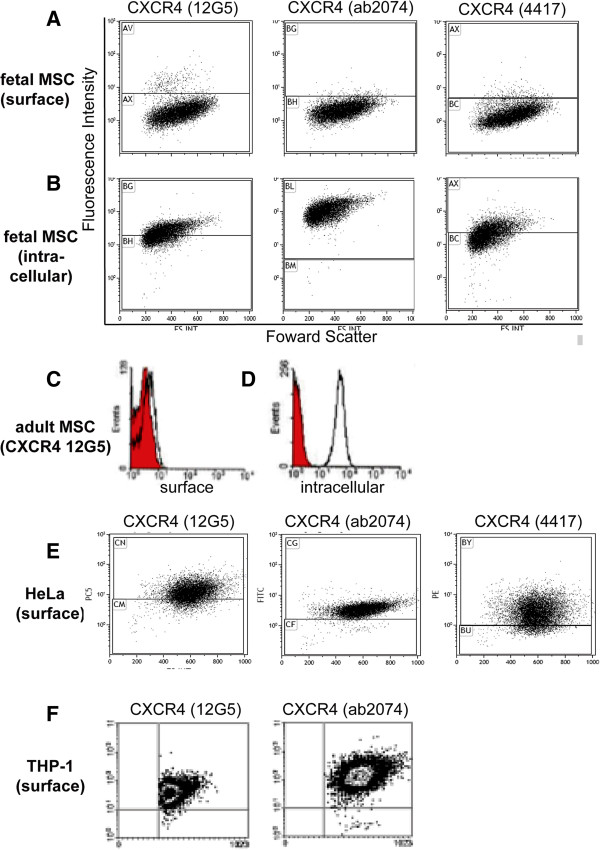
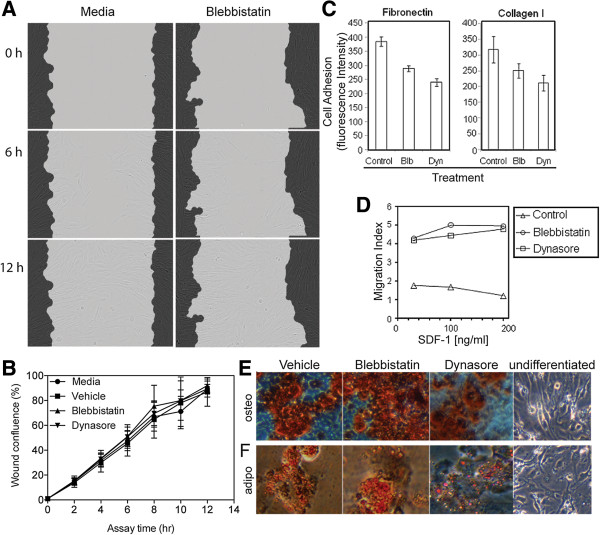
Results: We found that up to 4% of human fetal MSC have detectable surface-localized CXCR4. In the majority of cells, CXCR4 is located not at the cell surface, as would be required for 'sensing' migratory cues, but intracellularly. CXCR4 was identified in early endosomes, recycling endosomes, and lysosomes, indicating only a small percentage of CXCR4 travelling to the plasma membrane. Notably CXCR4 was also found in and around the nucleus, as detected with an anti-CXCR4 antibody directed specifically against CXCR4 isoform 2 differing only in N-terminal sequence. After demonstrating that endocytosis of CXCR4 is largely independent of endogenously-produced SDF-1, we next applied the cytoskeletal inhibitors blebbistatin and dynasore to inhibit endocytotic recycling. These increased the number of cells expressing surface CXCR4 by 10 and 5 fold respectively, and enhanced the number of cells migrating to SDF1 in vitro (up to 2.6 fold). These molecules had a transient effect on cell morphology and adhesion, which abated after the removal of the inhibitors, and did not alter functional stem cell properties.
Conclusions: We conclude that constitutive endocytosis is implicated in the regulation of CXCR4 membrane expression, and suggest a novel pharmacological strategy to enhance migration of systemically-transplanted cells.
Figures




Similar articles
-
[Role of endocytosis in cell surface CXC chemokine receptor 4 expression of stem cells from apical papilla].Beijing Da Xue Xue Bao Yi Xue Ban. 2019 Oct 18;51(5):893-899. doi: 10.19723/j.issn.1671-167X.2019.05.017. Beijing Da Xue Xue Bao Yi Xue Ban. 2019. PMID: 31624395 Free PMC article. Chinese.
-
Stromal cell-derived factor-1 receptor CXCR4-overexpressing bone marrow mesenchymal stem cells accelerate wound healing by migrating into skin injury areas.Cell Reprogram. 2013 Jun;15(3):206-15. doi: 10.1089/cell.2012.0046. Cell Reprogram. 2013. PMID: 23713431 Free PMC article.
-
Stromal cell-derived factor 1/CXCR4 signaling is critical for the recruitment of mesenchymal stem cells to the fracture site during skeletal repair in a mouse model.Arthritis Rheum. 2009 Mar;60(3):813-23. doi: 10.1002/art.24330. Arthritis Rheum. 2009. PMID: 19248097
-
Trafficking of normal stem cells and metastasis of cancer stem cells involve similar mechanisms: pivotal role of the SDF-1-CXCR4 axis.Stem Cells. 2005 Aug;23(7):879-94. doi: 10.1634/stemcells.2004-0342. Epub 2005 May 11. Stem Cells. 2005. PMID: 15888687 Review.
-
Mechanism of human stem cell migration and repopulation of NOD/SCID and B2mnull NOD/SCID mice. The role of SDF-1/CXCR4 interactions.Ann N Y Acad Sci. 2001 Jun;938:83-95. doi: 10.1111/j.1749-6632.2001.tb03577.x. Ann N Y Acad Sci. 2001. PMID: 11458529 Review.
Cited by
-
Isolation and Expansion of Mesenchymal Stem/Stromal Cells Derived from Human Placenta Tissue.J Vis Exp. 2016 Jun 6;(112):54204. doi: 10.3791/54204. J Vis Exp. 2016. PMID: 27340821 Free PMC article.
-
Adeno-Associated Virus Vectors and Stem Cells: Friends or Foes?Hum Gene Ther. 2017 Jun;28(6):450-463. doi: 10.1089/hum.2017.038. Hum Gene Ther. 2017. PMID: 28490211 Free PMC article. Review.
-
Transcriptional ontogeny of first trimester human fetal and placental mesenchymal stem cells: Gestational age versus niche.Genom Data. 2014 Oct 25;2:382-5. doi: 10.1016/j.gdata.2014.10.016. eCollection 2014 Dec. Genom Data. 2014. PMID: 26484134 Free PMC article.
-
Interaction of immune checkpoint PD-1 and chemokine receptor 4 (CXCR4) promotes a malignant phenotype in pancreatic cancer cells.PLoS One. 2022 Jul 7;17(7):e0270832. doi: 10.1371/journal.pone.0270832. eCollection 2022. PLoS One. 2022. PMID: 35797269 Free PMC article.
-
Impaired mRNA Expression of the Migration Related Chemokine Receptor CXCR4 in Mesenchymal Stem Cells of COPD Patients.Int J Inflam. 2017;2017:6089425. doi: 10.1155/2017/6089425. Epub 2017 Jul 19. Int J Inflam. 2017. PMID: 28804668 Free PMC article.
References
-
- Guillot PV, Gotherstrom C, Chan J, Kurata H, Fisk NM. Human first-trimester fetal MSC express pluripotency markers and grow faster and have longer telomeres than adult MSC. Stem Cells. 2007;25(3):646–654. - PubMed
-
- World’s first cell race no small affair. Updated [ http://blogs.nature.com/news/2011/12/worlds_first_cell_race_no_smal.html]
-
- Ramirez M, Lucia A, Gomez-Gallego F, Esteve-Lanao J, Perez-Martinez A, Foster C, Andreu AL, Martin MA, Madero L, Arenas J, García-Castro J. Mobilisation of mesenchymal cells into blood in response to skeletal muscle injury. Br J Sports Med. 2006;40(8):719–722. doi: 10.1136/bjsm.2006.028639. - DOI - PMC - PubMed
-
- Horita Y, Honmou O, Harada K, Houkin K, Hamada H, Kocsis JD. Intravenous administration of glial cell line-derived neurotrophic factor gene-modified human mesenchymal stem cells protects against injury in a cerebral ischemia model in the adult rat. J Neurosci Res. 2006;84(7):1495–1504. doi: 10.1002/jnr.21056. - DOI - PMC - PubMed
-
- Hauger O, Frost EE, van Heeswijk R, Deminiere C, Xue R, Delmas Y, Combe C, Moonen CT, Grenier N, Bulte JW. MR evaluation of the glomerular homing of magnetically labeled mesenchymal stem cells in a rat model of nephropathy. Radiology. 2006;238(1):200–210. doi: 10.1148/radiol.2381041668. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
