

Transcriptomic and metabolomic analysis of copper stress acclimation in Ectocarpus siliculosus highlights signaling and tolerance mechanisms in brown algae
- PMID: 24885189
- PMCID: PMC4108028
- DOI: 10.1186/1471-2229-14-116
Transcriptomic and metabolomic analysis of copper stress acclimation in Ectocarpus siliculosus highlights signaling and tolerance mechanisms in brown algae
Abstract
Background: Brown algae are sessile macro-organisms of great ecological relevance in coastal ecosystems. They evolved independently from land plants and other multicellular lineages, and therefore hold several original ontogenic and metabolic features. Most brown algae grow along the coastal zone where they face frequent environmental changes, including exposure to toxic levels of heavy metals such as copper (Cu).
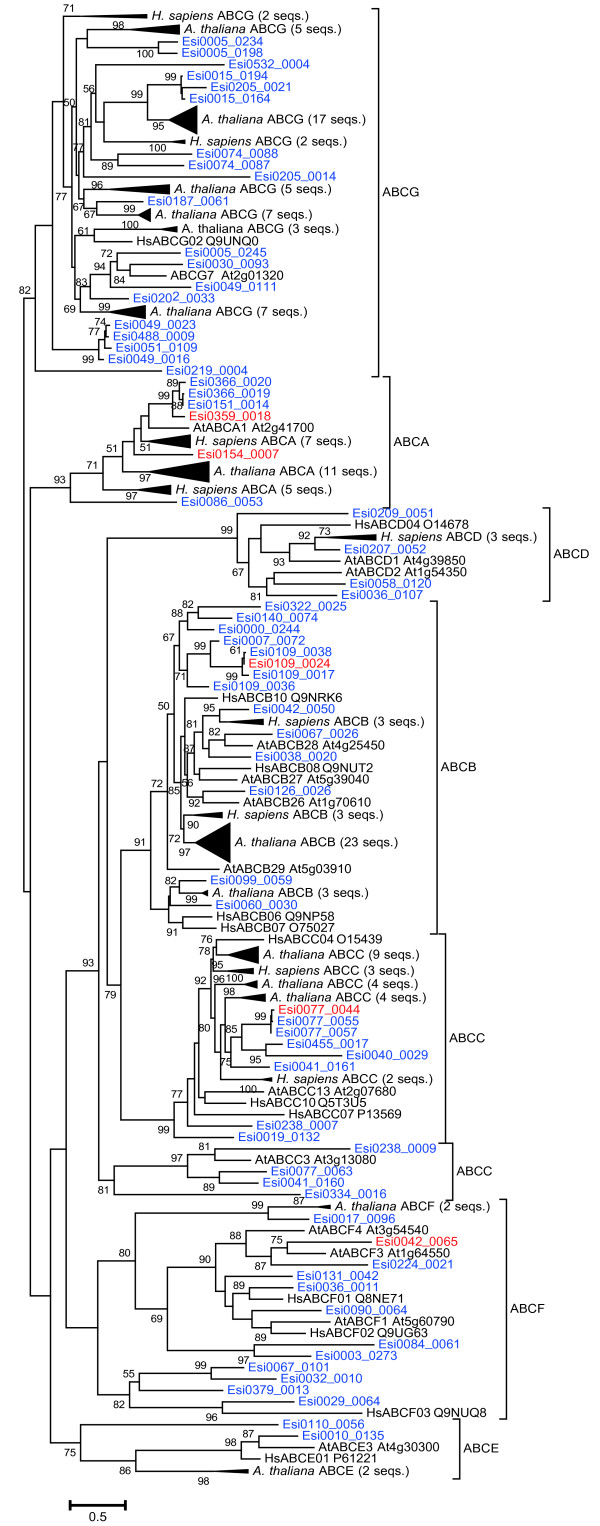
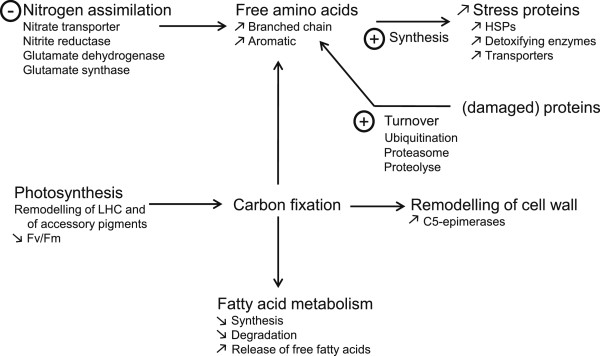
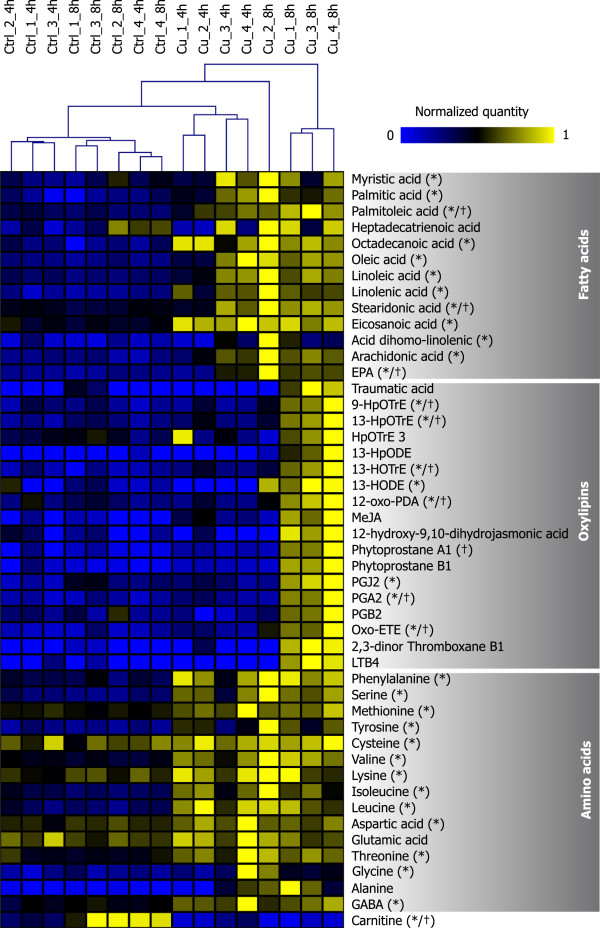
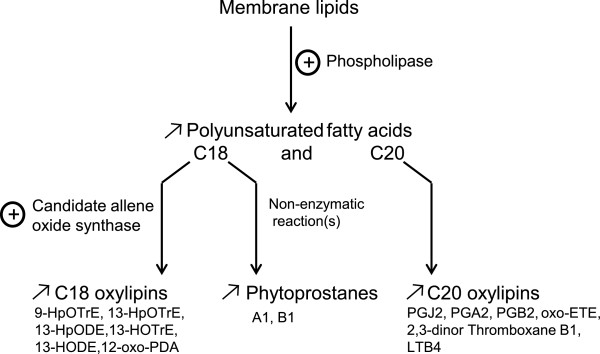
Results: We carried out large-scale transcriptomic and metabolomic analyses to decipher the short-term acclimation of the brown algal model E. siliculosus to Cu stress, and compared these data to results known for other abiotic stressors. This comparison demonstrates that Cu induces oxidative stress in E. siliculosus as illustrated by the transcriptomic overlap between Cu and H2O2 treatments. The common response to Cu and H2O2 consisted in the activation of the oxylipin and the repression of inositol signaling pathways, together with the regulation of genes coding for several transcription-associated proteins. Concomitantly, Cu stress specifically activated a set of genes coding for orthologs of ABC transporters, a P1B-type ATPase, ROS detoxification systems such as a vanadium-dependent bromoperoxidase, and induced an increase of free fatty acid contents. Finally we observed, as a common abiotic stress mechanism, the activation of autophagic processes on one hand and the repression of genes involved in nitrogen assimilation on the other hand.
Conclusions: Comparisons with data from green plants indicate that some processes involved in Cu and oxidative stress response are conserved across these two distant lineages. At the same time the high number of yet uncharacterized brown alga-specific genes induced in response to copper stress underlines the potential to discover new components and molecular interactions unique to these organisms. Of particular interest for future research is the potential cross-talk between reactive oxygen species (ROS)-, myo-inositol-, and oxylipin signaling.
Figures
Similar articles
-
Global expression analysis of the brown alga Ectocarpus siliculosus (Phaeophyceae) reveals large-scale reprogramming of the transcriptome in response to abiotic stress.Genome Biol. 2009;10(6):R66. doi: 10.1186/gb-2009-10-6-r66. Epub 2009 Jun 16. Genome Biol. 2009. PMID: 19531237 Free PMC article.
-
Integrative analysis of metabolite and transcript abundance during the short-term response to saline and oxidative stress in the brown alga Ectocarpus siliculosus.Plant Cell Environ. 2011 Apr;34(4):629-42. doi: 10.1111/j.1365-3040.2010.02268.x. Epub 2011 Feb 1. Plant Cell Environ. 2011. PMID: 21281312
-
Copper-induced intra-specific oxidative damage and antioxidant responses in strains of the brown alga Ectocarpus siliculosus with different pollution histories.Aquat Toxicol. 2015 Feb;159:81-9. doi: 10.1016/j.aquatox.2014.11.019. Epub 2014 Dec 2. Aquat Toxicol. 2015. PMID: 25521566
-
Toward systems biology in brown algae to explore acclimation and adaptation to the shore environment.OMICS. 2011 Dec;15(12):883-92. doi: 10.1089/omi.2011.0089. Epub 2011 Dec 2. OMICS. 2011. PMID: 22136637 Review.
-
Development and physiology of the brown alga Ectocarpus siliculosus: two centuries of research.New Phytol. 2008;177(2):319-332. doi: 10.1111/j.1469-8137.2007.02304.x. New Phytol. 2008. PMID: 18181960 Review.
Cited by
-
A Novel Protein from Ectocarpus sp. Improves Salinity and High Temperature Stress Tolerance in Arabidopsis thaliana.Int J Mol Sci. 2021 Feb 17;22(4):1971. doi: 10.3390/ijms22041971. Int J Mol Sci. 2021. PMID: 33671243 Free PMC article.
-
Biologically Active Oxylipins from Enzymatic and Nonenzymatic Routes in Macroalgae.Mar Drugs. 2016 Jan 20;14(1):23. doi: 10.3390/md14010023. Mar Drugs. 2016. PMID: 26805855 Free PMC article. Review.
-
Evolution and Expansion of the Prokaryote-Like Lipoxygenase Family in the Brown Alga Saccharina japonica.Front Plant Sci. 2017 Nov 28;8:2018. doi: 10.3389/fpls.2017.02018. eCollection 2017. Front Plant Sci. 2017. PMID: 29234336 Free PMC article.
-
Evolutionary conservation and metabolic significance of autophagy in algae.Philos Trans R Soc Lond B Biol Sci. 2024 Nov 18;379(1914):20230368. doi: 10.1098/rstb.2023.0368. Epub 2024 Sep 30. Philos Trans R Soc Lond B Biol Sci. 2024. PMID: 39343016 Free PMC article. Review.
-
Transcriptomic Analysis Reveals the Roles of Detoxification Systems in Response to Mercury in Chromera velia.Biomolecules. 2019 Oct 24;9(11):647. doi: 10.3390/biom9110647. Biomolecules. 2019. PMID: 31653042 Free PMC article.
References
-
- Correa JA, Castilla J, Ramirez M, Varas M, Lagos N, Vergara S, Moenne A, Roman D, Brown MT. Copper, copper mine tailings and their effect on the marine algae in Northern Chile. J Appl Phycol. 1999;11:57–67. doi: 10.1023/A:1008027610826. - DOI
-
- Gledhill M, Nimmo M, Hill SJ, Brown MT. The toxicity of copper (II) species to marine algae, with particular reference to macroalgae. J Phycol. 1997;33:2–11. doi: 10.1111/j.0022-3646.1997.00002.x. - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
