

Hippocampal neuronal cells that accumulate α-synuclein fragments are more vulnerable to Aβ oligomer toxicity via mGluR5--implications for dementia with Lewy bodies
- PMID: 24885390
- PMCID: PMC4041038
- DOI: 10.1186/1750-1326-9-18
Hippocampal neuronal cells that accumulate α-synuclein fragments are more vulnerable to Aβ oligomer toxicity via mGluR5--implications for dementia with Lewy bodies
Retraction in
-
Retraction Note: Hippocampal neuronal cells that accumulate α-synuclein fragments are more vulnerable to Aβ oligomer toxicity via mGluR5- implications for dementia with lewy bodies.Mol Neurodegener. 2025 Aug 5;20(1):87. doi: 10.1186/s13024-025-00881-6. Mol Neurodegener. 2025. PMID: 40759974 Free PMC article. No abstract available.
Abstract
Background: In dementia with Lewy bodies (DLB) abnormal interactions between α-synuclein (α-syn) and beta amyloid (Aβ) result in selective degeneration of neurons in the neocortex, limbic system and striatum. However, factors rendering these neurons selectively vulnerable have not been fully investigated. The metabotropic glutamate receptor 5 (mGluR5) has been shown to be up regulated in DLB and might play a role as a mediator of the neurotoxic effects of Aβ and α-syn in vulnerable neuronal populations. In this context, the main objective of the present study was to investigate the role of mGluR5 as a mediator of the neurotoxic effects of α-syn and Aβ in the hippocampus.
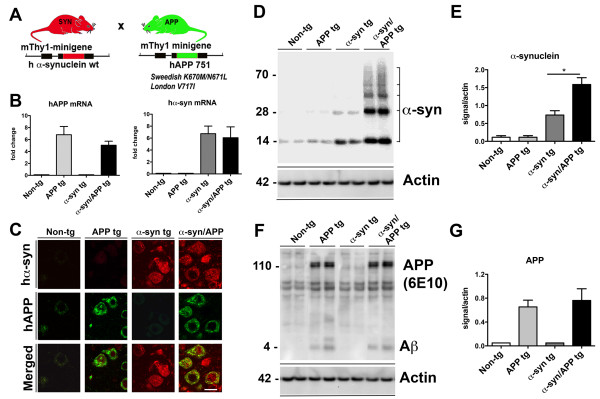
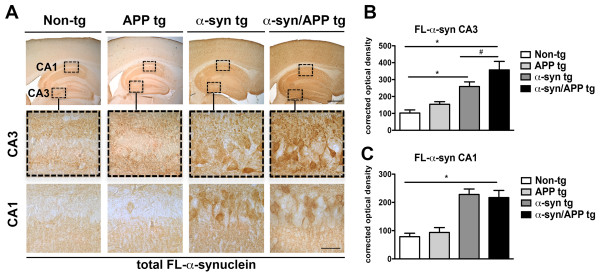
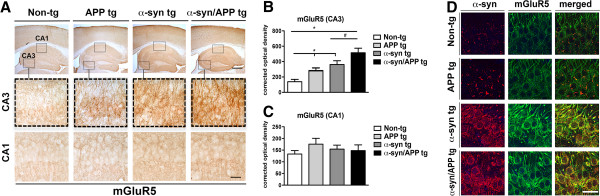
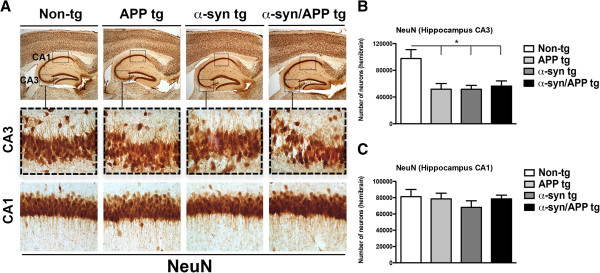
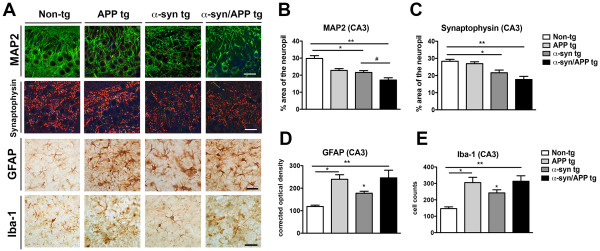
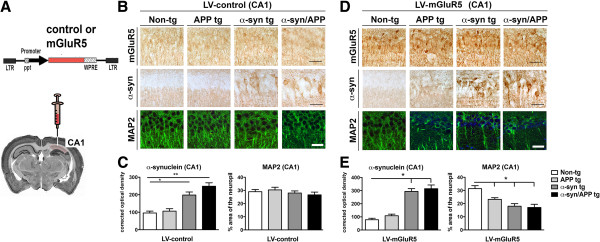
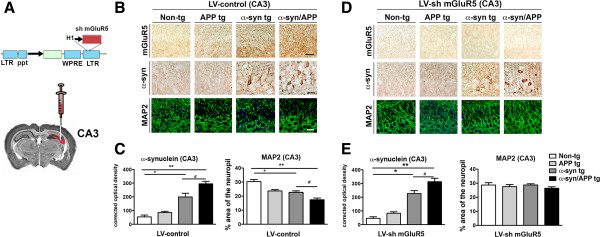
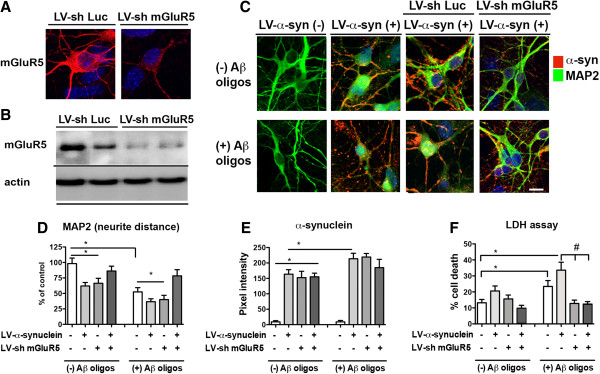
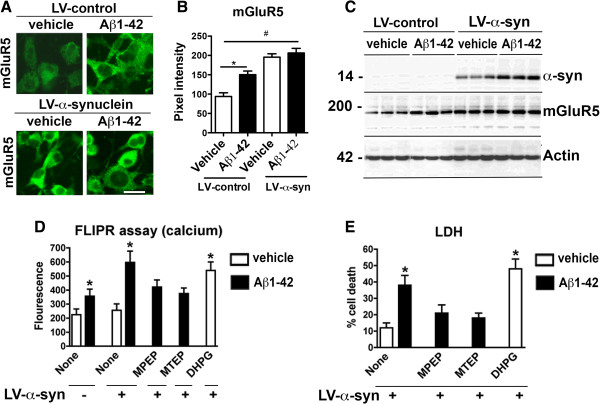
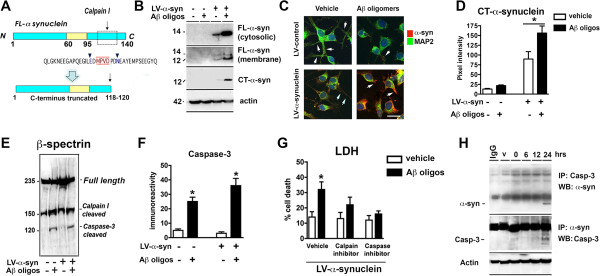
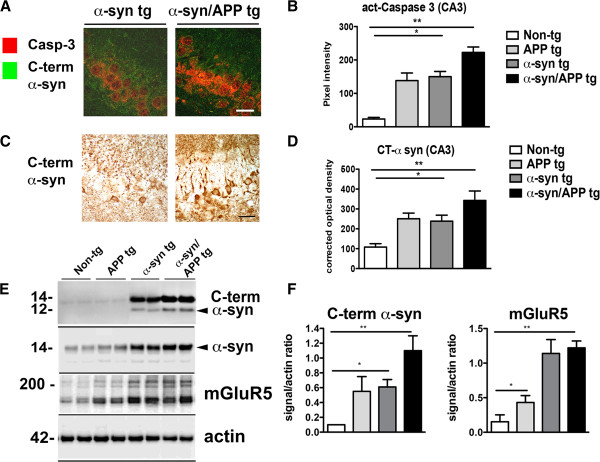
Results: We generated double transgenic mice over-expressing amyloid precursor protein (APP) and α-syn under the mThy1 cassette and investigated the relationship between α-syn cleavage, Aβ, mGluR5 and neurodegeneration in the hippocampus. We found that compared to the single tg mice, the α-syn/APP tg mice displayed greater accumulation of α-syn and mGluR5 in the CA3 region of the hippocampus compared to the CA1 and other regions. This was accompanied by loss of CA3 (but not CA1) neurons in the single and α-syn/APP tg mice and greater loss of MAP 2 and synaptophysin in the CA3 in the α-syn/APP tg. mGluR5 gene transfer using a lentiviral vector into the hippocampus CA1 region resulted in greater α-syn accumulation and neurodegeneration in the single and α-syn/APP tg mice. In contrast, silencing mGluR5 with a lenti-shRNA protected neurons in the CA3 region of tg mice. In vitro, greater toxicity was observed in primary hippocampal neuronal cultures treated with Aβ oligomers and over-expressing α-syn; this effect was attenuated by down-regulating mGluR5 with an shRNA lentiviral vector. In α-syn-expressing neuronal cells lines, Aβ oligomers promoted increased intracellular calcium levels, calpain activation and α-syn cleavage resulting in caspase-3-dependent cell death. Treatment with pharmacological mGluR5 inhibitors such as 2-Methyl-6-(phenylethynyl)pyridine (MPEP) and 3-((2-Methyl-4-thiazolyl)ethynyl)pyridine (MTEP) attenuated the toxic effects of Aβ in α-syn-expressing neuronal cells.
Conclusions: Together, these results support the possibility that vulnerability of hippocampal neurons to α-syn and Aβ might be mediated via mGluR5. Moreover, therapeutical interventions targeting mGluR5 might have a role in DLB.
Figures
References
-
- As A. Alzheimer’s disease facts and figures. Alzheimer’s Dementia. 2013;2013(9):1–71. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
