

The genetic architecture of constitutive and induced trichome density in two new recombinant inbred line populations of Arabidopsis thaliana: phenotypic plasticity, epistasis, and bidirectional leaf damage response
- PMID: 24885520
- PMCID: PMC4108038
- DOI: 10.1186/1471-2229-14-119
The genetic architecture of constitutive and induced trichome density in two new recombinant inbred line populations of Arabidopsis thaliana: phenotypic plasticity, epistasis, and bidirectional leaf damage response
Abstract
Background: Herbivory imposes an important selective pressure on plants. In Arabidopsis thaliana leaf trichomes provide a key defense against insect herbivory; however, trichome production incurs a fitness cost in the absence of herbivory. Previous work on A. thaliana has shown an increase in trichome density in response to leaf damage, suggesting a mechanism by which the cost associated with constitutively high trichome density might be mitigated; however, the genetic basis of trichome density induction has not been studied.
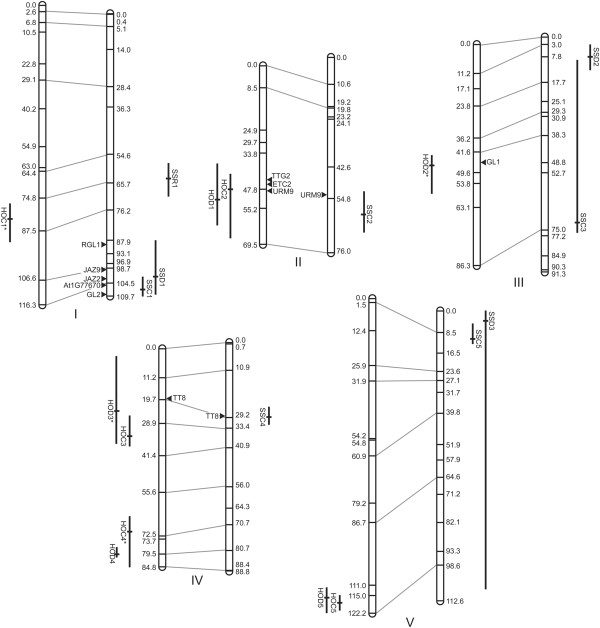
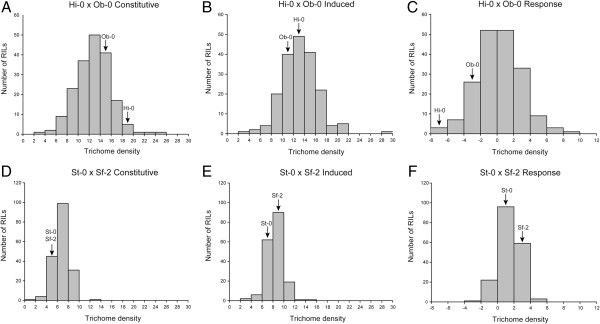
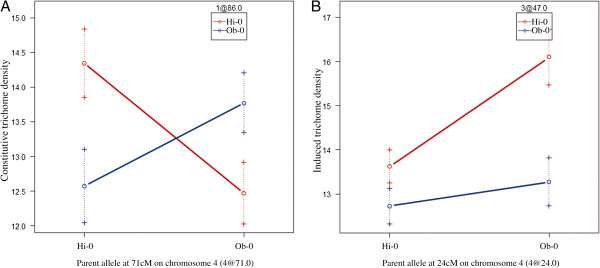
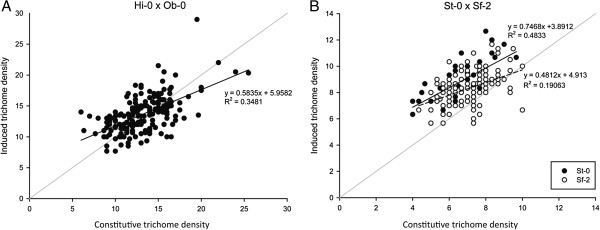
Results: Here, we describe the mapping of quantitative trait loci (QTL) for constitutive and damage induced trichome density in two new recombinant inbred line populations of A. thaliana; mapping for constitutive and induced trichome density also allowed for the investigation of damage response (plasticity) QTL. Both novel and previously identified QTL for constitutive trichome density and the first QTL for induced trichome density and response are identified. Interestingly, two of the four parental accessions and multiple RILs in each population exhibited lower trichome density following leaf damage, a response not previously described in A. thaliana. Importantly, a single QTL was mapped for the response phenotype and allelic variation at this locus appears to determine response trajectory in RILs. The data also show that epistatic interactions are a significant component of the genetic architecture of trichome density.
Conclusions: Together, our results provide further insights into the genetic architecture of constitutive trichome density and new insights into induced trichome density in A. thaliana specifically and to our understanding of the genetic underpinnings of natural variation generally.
Figures
References
-
- Cyr H, Pace ML. Magnitude and patterns of herbivory in aquatic and terrestrial ecosystems. Nature. 1993;361(6408):148–150. doi: 10.1038/361148a0. - DOI
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
