

Adaptive and pathological inhibition of neuroplasticity associated with circadian rhythms and sleep
- PMID: 24886189
- PMCID: PMC4060045
- DOI: 10.1037/a0036689
Adaptive and pathological inhibition of neuroplasticity associated with circadian rhythms and sleep
Abstract
The circadian system organizes sleep and wake through imposing a daily cycle of sleep propensity on the organism. Sleep has been shown to play an important role in learning and memory. Apart from the daily cycle of sleep propensity, however, direct effects of the circadian system on learning and memory also have been well documented. Many mechanistic components of the memory consolidation process ranging from the molecular to the systems level have been identified and studied. The question that remains is how do these various processes and components work together to produce cycles of increased and decreased learning abilities, and why should there be times of day when neural plasticity appears to be restricted? Insights into this complex problem can be gained through investigations of the learning disabilities caused by circadian disruption in Siberian hamsters and by aneuploidy in Down's syndrome mice. A simple working hypothesis that has been explored in this work is that the observed learning disabilities are due to an altered excitation/inhibition balance in the CNS. Excessive inhibition is the suspected cause of deficits in memory consolidation. In this article we present the evidence that excessive inhibition in these cases of learning disability involves GABAergic neurotransmission, that treatment with GABA receptor inhibitors can reverse the learning disability, and that the efficacy of the treatment is time sensitive coincident with the major daily sleep phase, and that it depends on sleep. The evidence we present leads us to hypothesize that a function of the circadian system is to reduce neuroplasticity during the daily sleep phase when processes of memory consolidation are taking place.
PsycINFO Database Record (c) 2014 APA, all rights reserved.
Figures


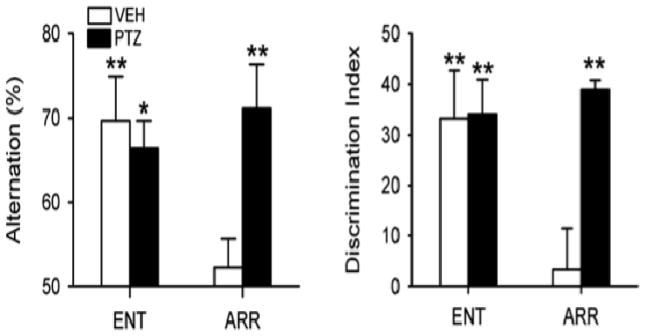


References
-
- Benington J, Heller HC. Restoration of brain energy metabolism as the function of sleep. Prog in Neurobiol. 1995;45:347–360. - PubMed
-
- Borbely A, Achermann P. Concepts and models of sleep regulation: an overview. Journal of Sleep Research. 1992;1:63–79. - PubMed
-
- Braudeau J, Delatour B, Duchon A, Pereira PL, Dauphinot L, de Chaumont F, Olivo-Marin JC, Dodd RH, Hérault Y, Potier MC. Specific targeting of the GABA-A receptor α5 subtype by a selective inverse agonist restores cognitive deficits in Down syndrome mice. J Psychopharmacol. 2011;25(8):1030–42. - PMC - PubMed
-
- Cain SW, Chou T, Ralph MR. Circadian modulation of performance on an aversion-based place learning task in hamsters. Behav Brain Res. 2004;150:201–205. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
