

DUSP3/VHR is a pro-angiogenic atypical dual-specificity phosphatase
- PMID: 24886454
- PMCID: PMC4038117
- DOI: 10.1186/1476-4598-13-108
DUSP3/VHR is a pro-angiogenic atypical dual-specificity phosphatase
Abstract
Background: DUSP3 phosphatase, also known as Vaccinia-H1 Related (VHR) phosphatase, encoded by DUSP3/Dusp3 gene, is a relatively small member of the dual-specificity protein phosphatases. In vitro studies showed that DUSP3 is a negative regulator of ERK and JNK pathways in several cell lines. On the other hand, DUSP3 is implicated in human cancer. It has been alternatively described as having tumor suppressive and oncogenic properties. Thus, the available data suggest that DUSP3 plays complex and contradictory roles in tumorigenesis that could be cell type-dependent. Since most of these studies were performed using recombinant proteins or in cell-transfection based assays, the physiological function of DUSP3 has remained elusive.
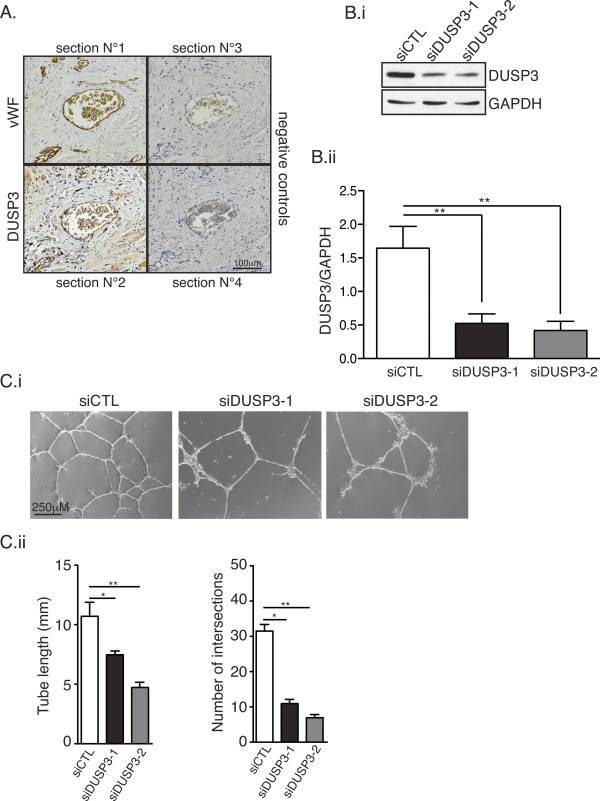
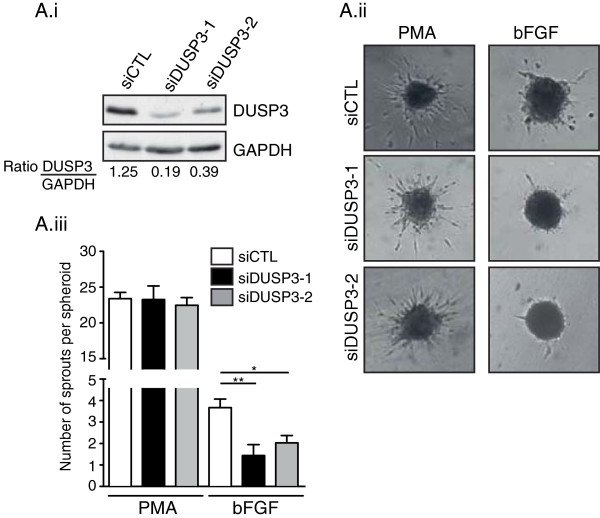
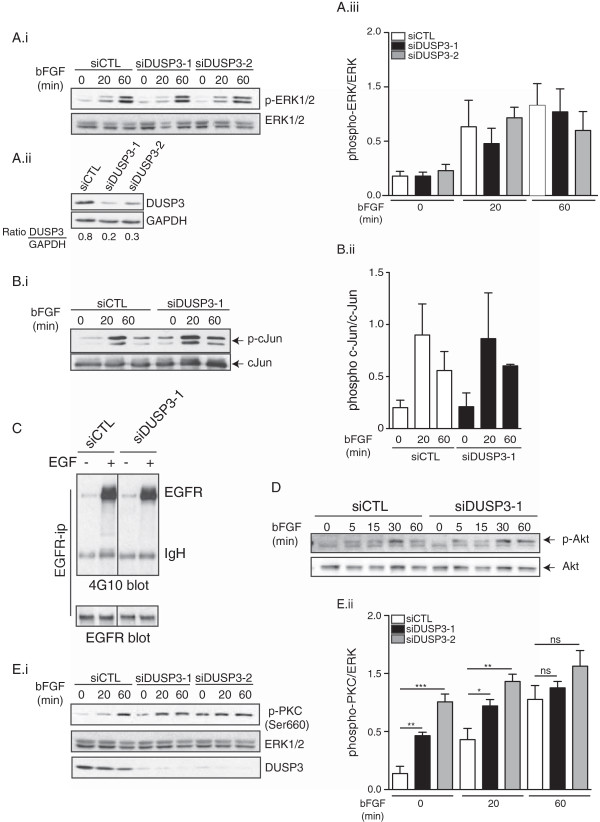
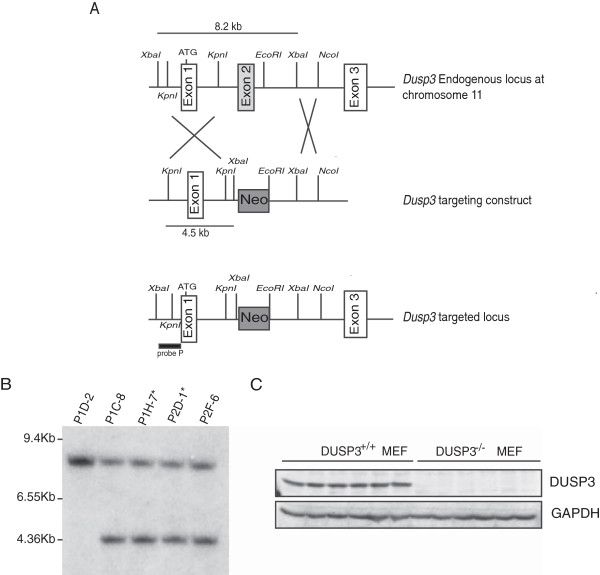
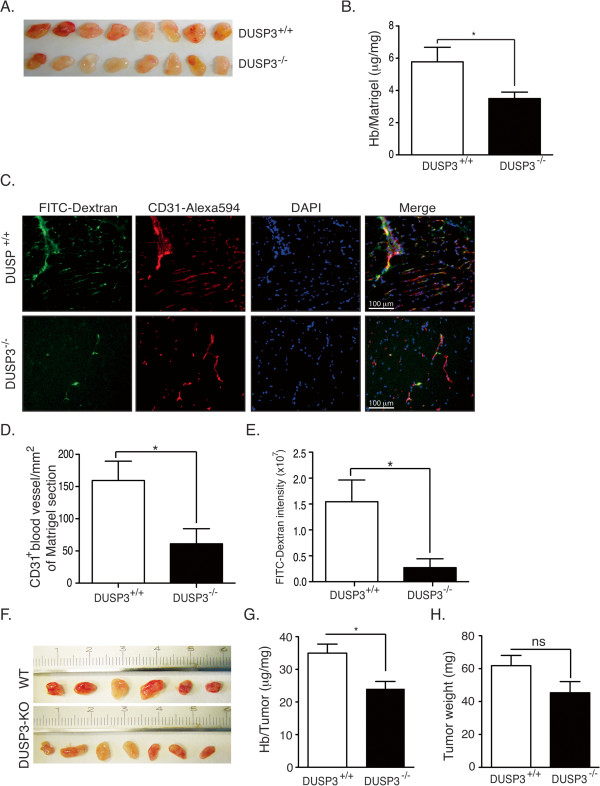
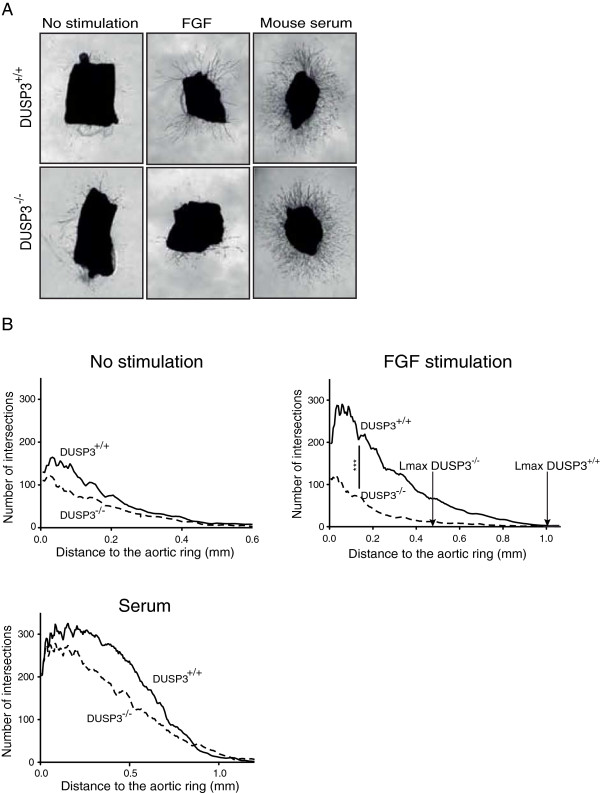
Results: Using immunohistochemistry on human cervical sections, we observed a strong expression of DUSP3 in endothelial cells (EC) suggesting a contribution for this phosphatase to EC functions. DUSP3 downregulation, using RNA interference, in human EC reduced significantly in vitro tube formation on Matrigel and spheroid angiogenic sprouting. However, this defect was not associated with an altered phosphorylation of the documented in vitro DUSP3 substrates, ERK1/2, JNK1/2 and EGFR but was associated with an increased PKC phosphorylation. To investigate the physiological function of DUSP3, we generated Dusp3-deficient mice by homologous recombination. The obtained DUSP3-/- mice were healthy, fertile, with no spontaneous phenotype and no vascular defect. However, DUSP3 deficiency prevented neo-vascularization of transplanted b-FGF containing Matrigel and LLC xenograft tumors as evidenced by hemoglobin (Hb) and FITC-dextran quantifications. Furthermore, we found that DUSP3 is required for b-FGF-induced microvessel outgrowth in the aortic ring assay.
Conclusions: All together, our data identify DUSP3 as a new important player in angiogenesis.
Figures

References
-
- Patterson KI, Brummer T, O’Brien PM, Daly RJ. Dual-specificity phosphatases: critical regulators with diverse cellular targets. Biochem J. 2009;418:475–489. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
