

G1 checkpoint establishment in vivo during embryonic liver development
- PMID: 24886500
- PMCID: PMC4031160
- DOI: 10.1186/1471-213X-14-23
G1 checkpoint establishment in vivo during embryonic liver development
Abstract
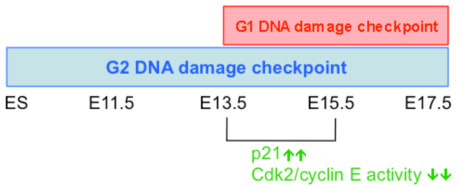
Background: The DNA damage-mediated cell cycle checkpoint is an essential mechanism in the DNA damage response (DDR). During embryonic development, the characteristics of cell cycle and DNA damage checkpoint evolve from an extremely short G1 cell phase and lacking G1 checkpoint to lengthening G1 phase and the establishment of the G1 checkpoint. However, the regulatory mechanisms governing these transitions are not well understood. In this study, pregnant mice were exposed to ionizing radiation (IR) to induce DNA damage at different embryonic stages; the kinetics and mechanisms of the establishment of DNA damage-mediated G1 checkpoint in embryonic liver were investigated.
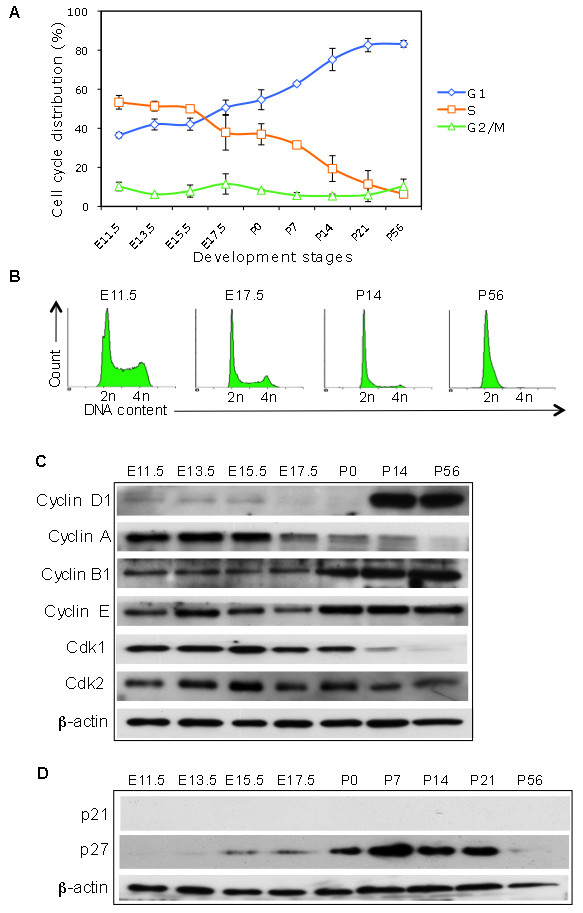
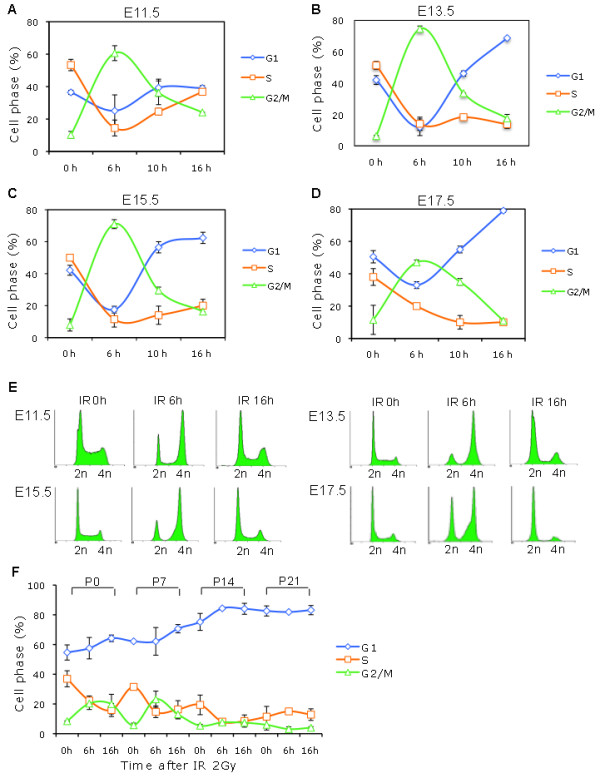
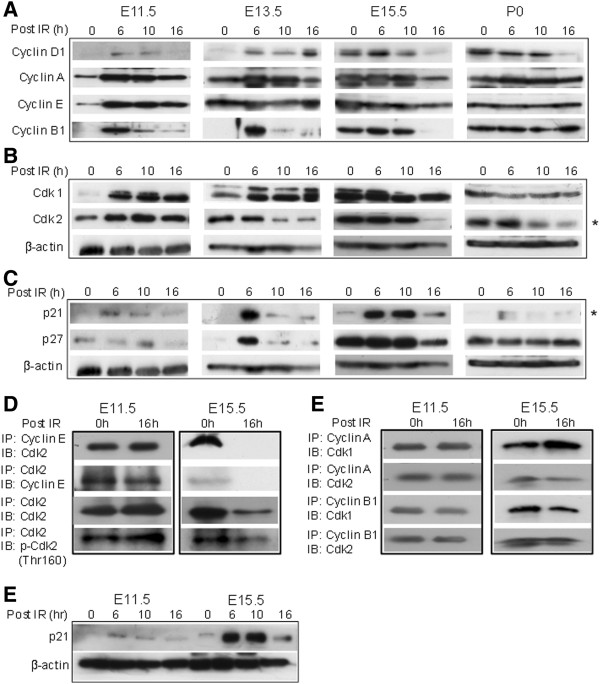
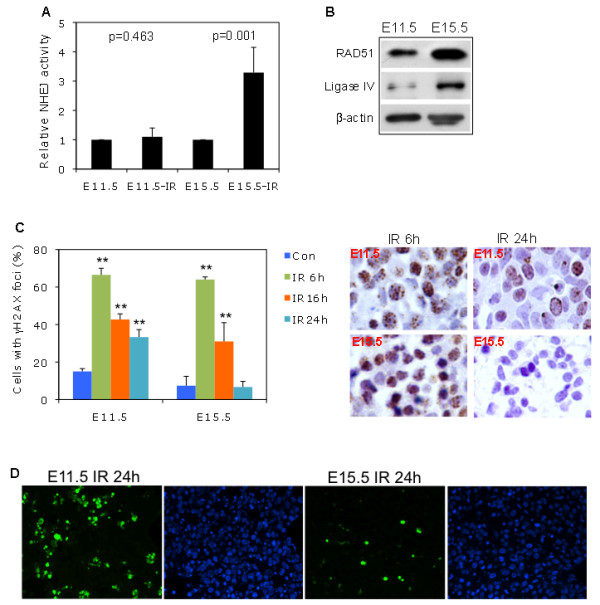
Results: We found that the G2 cell cycle arrest was the first response to DNA damage in early developmental stages. Starting at E13.5/E15.5, IR mediated inhibition of the G1 to S phase transition became evident. Concomitantly, IR induced the robust expression of p21 and suppressed Cdk2/cyclin E activity, which might involve in the initiation of G1 checkpoint. The established G1 cell cycle checkpoint, in combination with an enhanced DNA repair capacity at E15.5, displayed biologically protective effects of repairing DNA double-strand breaks (DSBs) and reducing apoptosis in the short term as well as reducing chromosome deletion and breakage in the long term.
Conclusion: Our study is the first to demonstrate the establishment of the DNA damage-mediated G1 cell cycle checkpoint in liver cells during embryogenesis and its in vivo biological effects during embryonic liver development.
Figures

Similar articles
-
Shift in G1-Checkpoint from ATM-Alone to a Cooperative ATM Plus ATR Regulation with Increasing Dose of Radiation.Cells. 2021 Dec 27;11(1):63. doi: 10.3390/cells11010063. Cells. 2021. PMID: 35011623 Free PMC article.
-
Lack of a p21waf1/cip -dependent G1/S checkpoint in neural stem and progenitor cells after DNA damage in vivo.Stem Cells. 2012 Mar;30(3):537-47. doi: 10.1002/stem.1010. Stem Cells. 2012. PMID: 22162343 Free PMC article.
-
An important role for CDK2 in G1 to S checkpoint activation and DNA damage response in human embryonic stem cells.Stem Cells. 2011 Apr;29(4):651-9. doi: 10.1002/stem.620. Stem Cells. 2011. PMID: 21319273
-
Cell cycle checkpoints and DNA repair preserve the stability of the human genome.Cancer Metastasis Rev. 1995 Mar;14(1):31-41. doi: 10.1007/BF00690209. Cancer Metastasis Rev. 1995. PMID: 7606819 Review.
-
[Cell cycle regulation after exposure to ionizing radiation].Bull Cancer. 1999 Apr;86(4):345-57. Bull Cancer. 1999. PMID: 10341340 Review. French.
Cited by
-
Sexually dimorphic DNA damage responses and mutation avoidance in the mouse germline.Genes Dev. 2020 Dec 1;34(23-24):1637-1649. doi: 10.1101/gad.341602.120. Epub 2020 Nov 12. Genes Dev. 2020. PMID: 33184219 Free PMC article.
-
XLF-mediated NHEJ activity in hepatocellular carcinoma therapy resistance.BMC Cancer. 2017 May 19;17(1):344. doi: 10.1186/s12885-017-3345-y. BMC Cancer. 2017. PMID: 28526069 Free PMC article.
References
-
- Milyavsky M, Gan OI, Trottier M, Komosa M, Tabach O, Notta F, Lechman E, Hermans KG, Eppert K, Konovalova Z, Omatsky O, Domany E, Meyn MS, Dick JE. A distinctive DNA damage response in human hematopoietic stem cells reveals an apoptosis-independent role for p53 in self-renewal. Cell Stem Cell. 2010;7:186–197. doi: 10.1016/j.stem.2010.05.016. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
