

Mechanisms of blindness: animal models provide insight into distinct CRX-associated retinopathies
- PMID: 24888636
- PMCID: PMC4177283
- DOI: 10.1002/dvdy.24151
Mechanisms of blindness: animal models provide insight into distinct CRX-associated retinopathies
Abstract
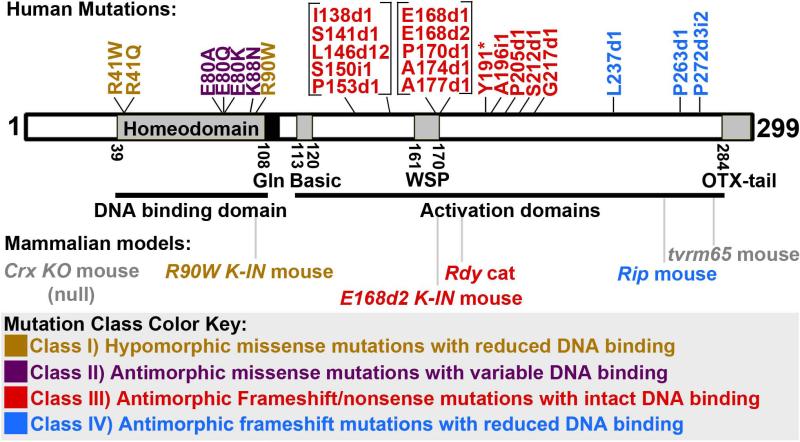
Background: The homeodomain transcription factor CRX is a crucial regulator of mammalian photoreceptor gene expression. Mutations in the human CRX gene are associated with dominant inherited retinopathies Retinitis Pigmentosa (RP), Cone-Rod Dystrophy (CoRD), and Leber Congenital Amaurosis (LCA), of varying severity. In vitro and in vivo assessment of mutant CRX proteins have revealed pathogenic mechanisms for several mutations, but no comprehensive mutation-disease correlation has yet been reported.
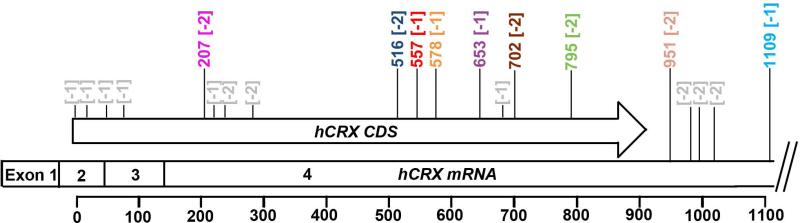
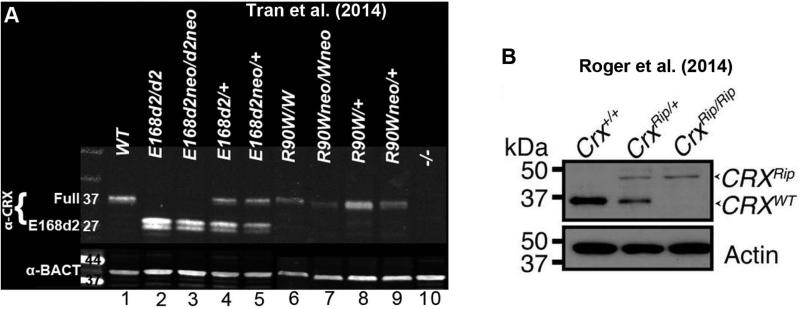
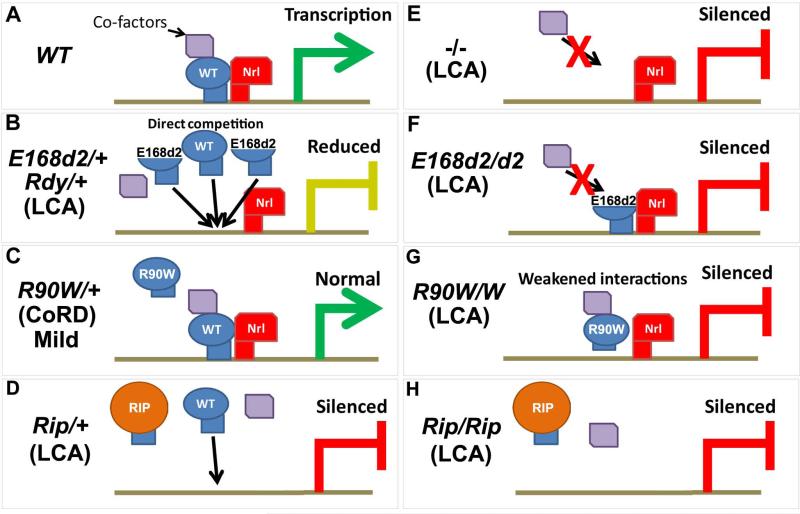
Results: Here we describe four different classes of disease-causing CRX mutations, characterized by mutation type, pathogenetic mechanism, and the molecular activity of the mutant protein: (1) hypomorphic missense mutations with reduced DNA binding, (2) antimorphic missense mutations with variable DNA binding, (3) antimorphic frameshift/nonsense mutations with intact DNA binding, and (4) antimorphic frameshift mutations with reduced DNA binding. Mammalian models representing three of these classes have been characterized.
Conclusions: Models carrying Class I mutations display a mild dominant retinal phenotype and recessive LCA, while models carrying Class III and IV mutations display characteristically distinct dominant LCA phenotypes. These animal models also reveal unexpected pathogenic mechanisms underlying CRX-associated retinopathies. The complexity of genotype-phenotype correlation for CRX-associated diseases highlights the value of developing comprehensive "true-to-disease" animal models for understanding pathologic mechanisms and testing novel therapeutic approaches.
Keywords: antimorph; disease models; dominant-negative; gene expression; human genetics; hypomorph; neural development; neuronal degeneration; photoreceptors; retina; transcription factors.
© 2014 Wiley Periodicals, Inc.
Figures
Similar articles
-
Mechanistically distinct mouse models for CRX-associated retinopathy.PLoS Genet. 2014 Feb 6;10(2):e1004111. doi: 10.1371/journal.pgen.1004111. eCollection 2014 Feb. PLoS Genet. 2014. PMID: 24516401 Free PMC article.
-
Retinopathy and attenuated circadian entrainment in Crx-deficient mice.Nat Genet. 1999 Dec;23(4):466-70. doi: 10.1038/70591. Nat Genet. 1999. PMID: 10581037
-
OTX2 loss causes rod differentiation defect in CRX-associated congenital blindness.J Clin Invest. 2014 Feb;124(2):631-43. doi: 10.1172/JCI72722. Epub 2014 Jan 2. J Clin Invest. 2014. PMID: 24382353 Free PMC article.
-
Dominant Leber congenital amaurosis, cone-rod degeneration, and retinitis pigmentosa caused by mutant versions of the transcription factor CRX.Hum Mutat. 2001 Dec;18(6):488-98. doi: 10.1002/humu.1226. Hum Mutat. 2001. PMID: 11748842 Review.
-
Gene Augmentation for Autosomal Dominant CRX-Associated Retinopathies.Adv Exp Med Biol. 2023;1415:135-141. doi: 10.1007/978-3-031-27681-1_21. Adv Exp Med Biol. 2023. PMID: 37440026 Free PMC article. Review.
Cited by
-
The microphthalmia-associated transcription factor (Mitf) gene and its role in regulating eye function.Sci Rep. 2019 Oct 28;9(1):15386. doi: 10.1038/s41598-019-51819-0. Sci Rep. 2019. PMID: 31659211 Free PMC article.
-
CRX directs photoreceptor differentiation by accelerating chromatin remodeling at specific target sites.Epigenetics Chromatin. 2018 Aug 1;11(1):42. doi: 10.1186/s13072-018-0212-2. Epigenetics Chromatin. 2018. PMID: 30068366 Free PMC article.
-
Genetic and epigenetic control of retinal development in zebrafish.Curr Opin Neurobiol. 2019 Dec;59:120-127. doi: 10.1016/j.conb.2019.05.008. Epub 2019 Jun 27. Curr Opin Neurobiol. 2019. PMID: 31255843 Free PMC article. Review.
-
Transcriptional precision in photoreceptor development and diseases - Lessons from 25 years of CRX research.Front Cell Neurosci. 2024 Feb 13;18:1347436. doi: 10.3389/fncel.2024.1347436. eCollection 2024. Front Cell Neurosci. 2024. PMID: 38414750 Free PMC article. Review.
-
Mutational scanning of CRX classifies clinical variants and reveals biochemical properties of the transcriptional effector domain.bioRxiv [Preprint]. 2024 Mar 27:2024.03.21.585809. doi: 10.1101/2024.03.21.585809. bioRxiv. 2024. Update in: Genome Res. 2024 Oct 29;34(10):1540-1552. doi: 10.1101/gr.279415.124. PMID: 38585983 Free PMC article. Updated. Preprint.
References
-
- Acland GM, Aguirre GD, Ray J, Zhang Q, Aleman TS, Cideciyan AV, Pearce-Kelling SE, Anand V, Zeng Y, Maguire AM, Jacobson SG, Hauswirth WW, Bennett J. Gene therapy restores vision in a canine model of childhood blindness. Nat Genet. 2001;28:92–95. - PubMed
-
- Baas D, Bumsted KM, Martinez JA, Vaccarino FM, Wikler KC, Barnstable CJ. The subcellular localization of Otx2 is cell-type specific and developmentally regulated in the mouse retina. Brain Res Mol Brain Res. 2000;78:26–37. - PubMed
-
- Bennicelli J, Wright JF, Komaromy A, Jacobs JB, Hauck B, Zelenaia O, Mingozzi F, Hui D, Chung D, Rex TS, Wei Z, Qu G, Zhou S, Zeiss C, Arruda VR, Acland GM, Dell'Osso LF, High KA, Maguire AM, Bennett J. Reversal of blindness in animal models of leber congenital amaurosis using optimized AAV2-mediated gene transfer. Mol Ther. 2008;16:458–465. - PMC - PubMed
-
- Berger W, Kloeckener-Gruissem B, Neidhardt J. The molecular basis of human retinal and vitreoretinal diseases. Progress in retinal and eye research. 2010;29(5):335–75. - PubMed
-
- Blackshaw S, Fraioli RE, Furukawa T, Cepko CL. Comprehensive analysis of photoreceptor gene expression and the identification of candidate retinal disease genes. Cell. 2001;107:579–589. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
