

Naturally occurring mutants inform SHBG structure and function
- PMID: 24892637
- PMCID: PMC5414826
- DOI: 10.1210/me.2014-1058
Naturally occurring mutants inform SHBG structure and function
Abstract
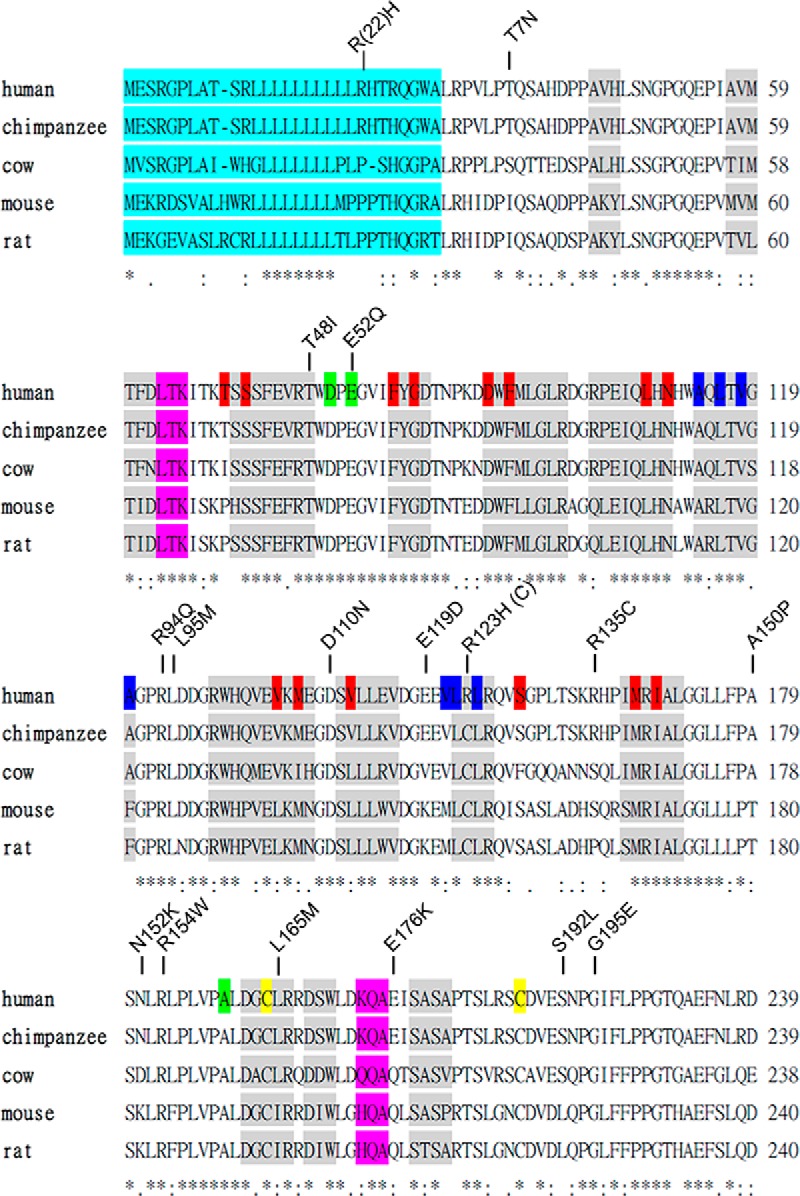
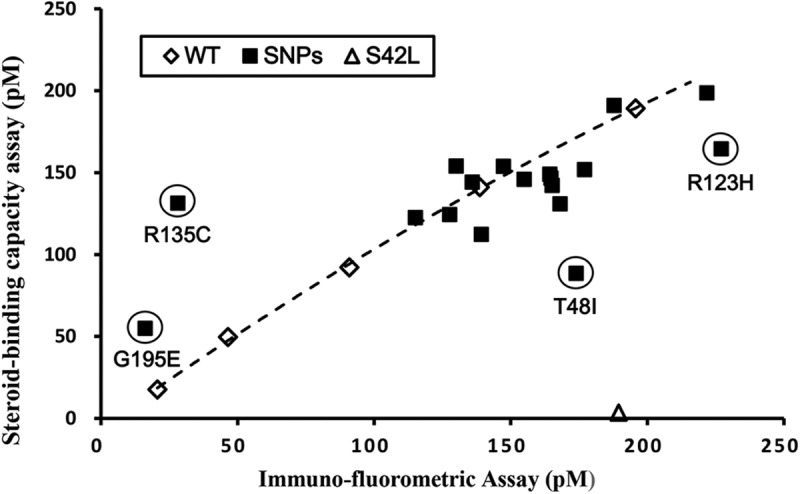
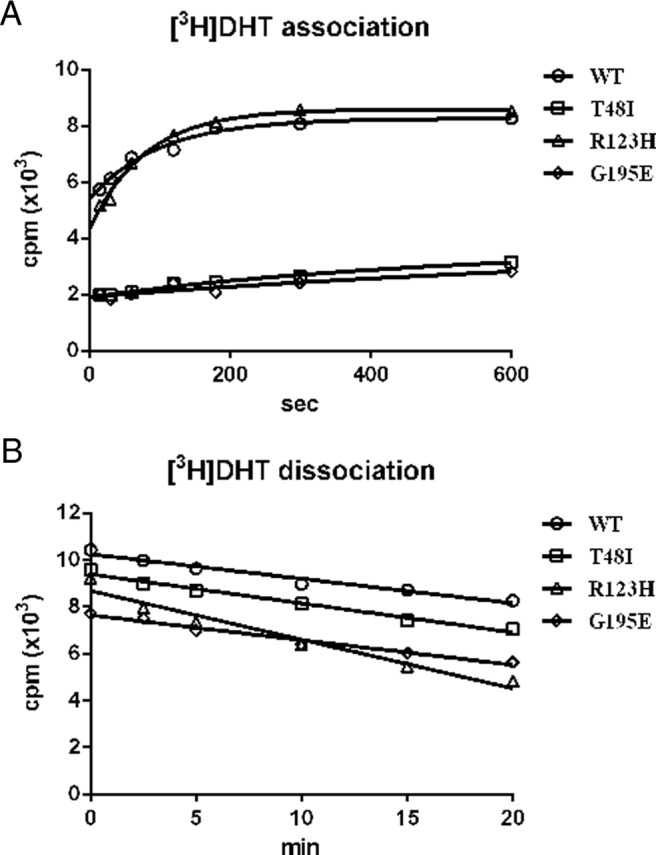
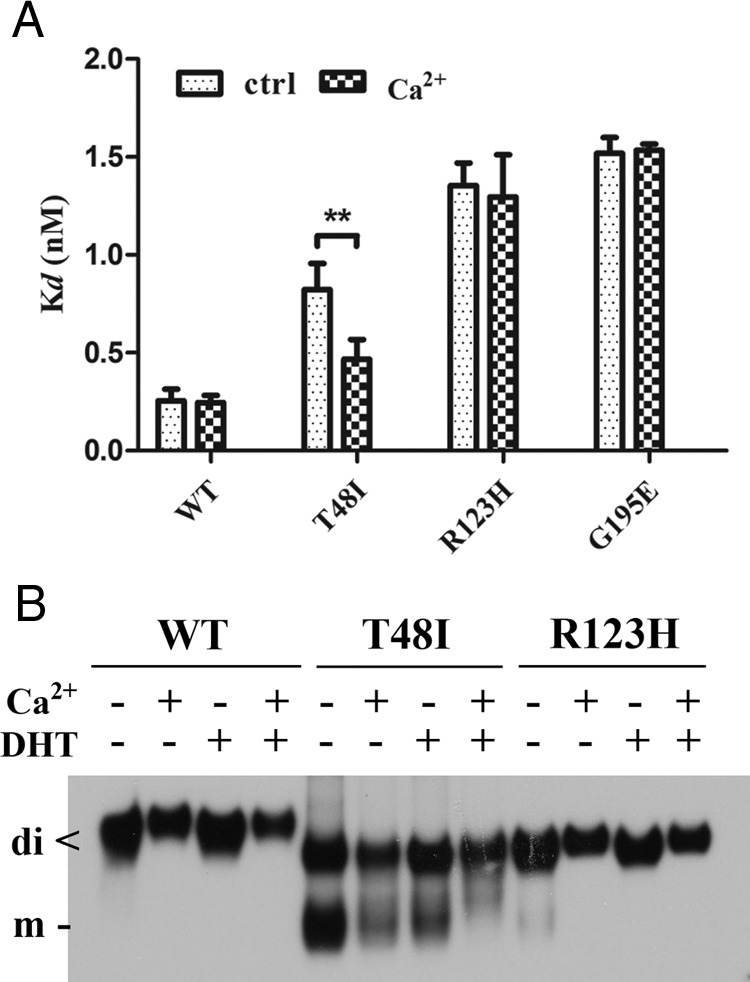
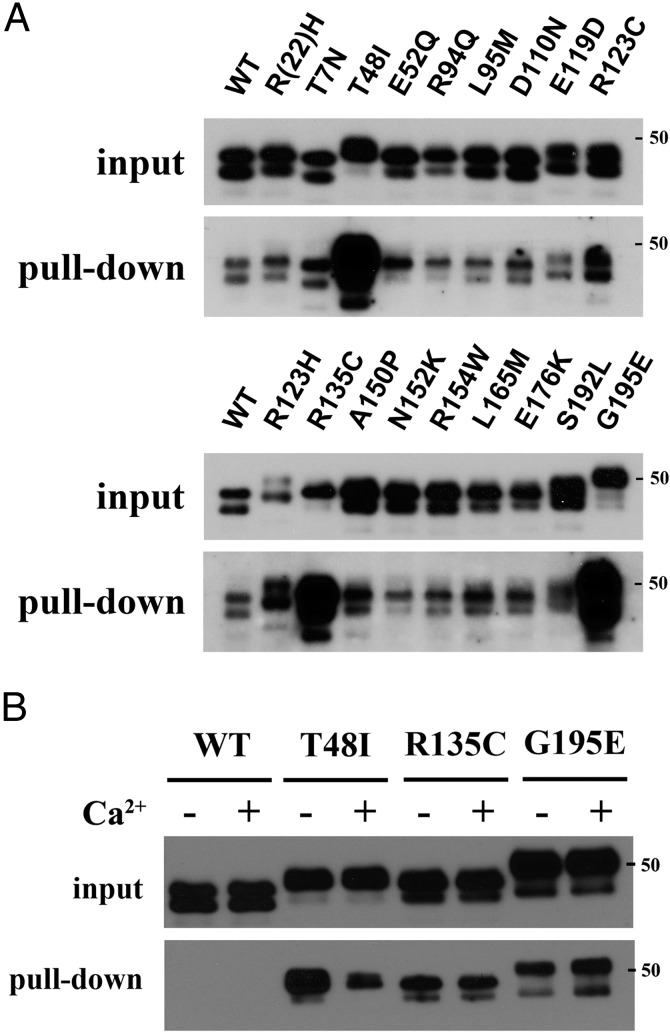
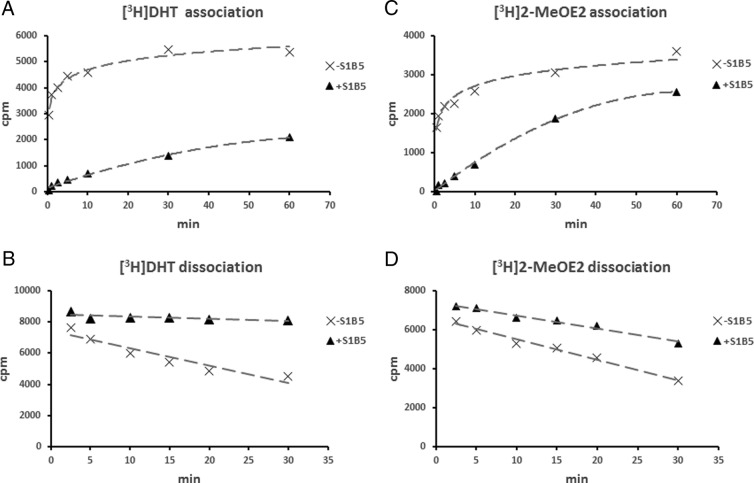
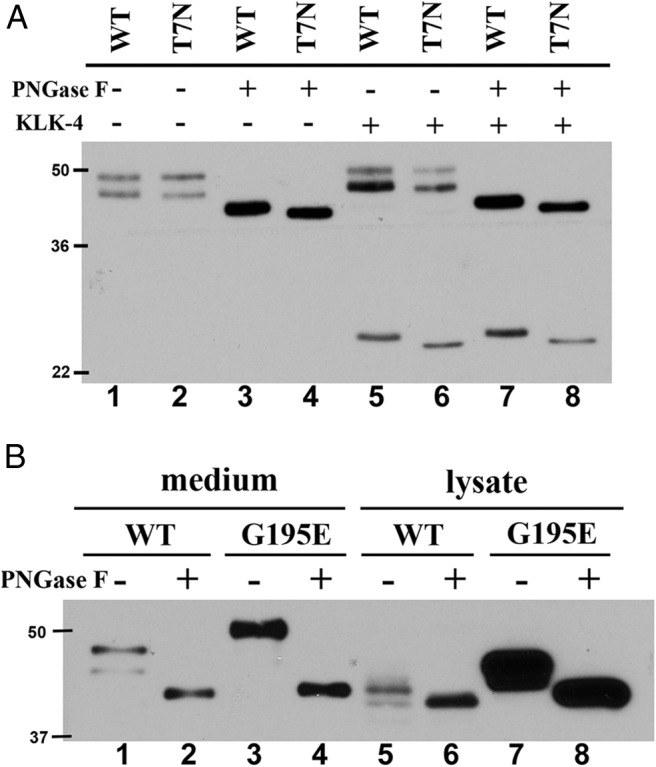
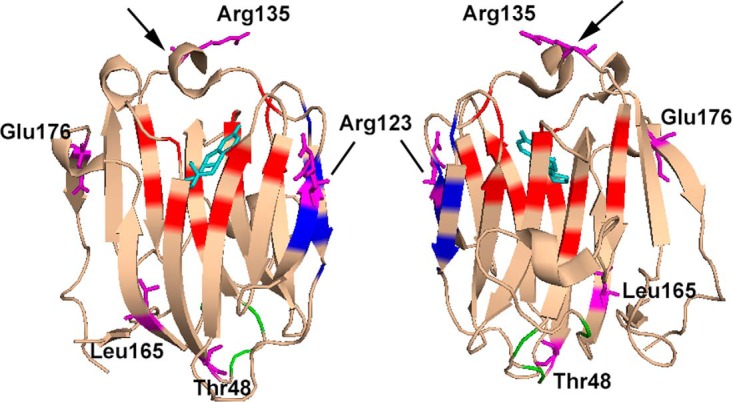
SHBG transports and regulates the activities of androgens and estrogens. Several single nucleotide polymorphisms in the human SHBG gene have been linked to sex steroid-dependent diseases, including those associated with the metabolic syndrome. The N-terminal laminin G-like domain of SHBG includes binding sites for calcium, sex steroids, and fibulin family members, as well as a dimerization domain. We have found that 8 of 18 uncharacterized nonsynonymous single nucleotide polymorphisms within this domain alter the production or biochemical properties of SHBG in ways not previously recognized. O-Linked glycosylation at Thr7 is disrupted in SHBG T7N, whereas abnormal glycosylation of SHBG G195E limits its secretion. Three SHBG mutants (R135C, L165M, and E176K) bind estradiol with abnormally high affinity. SHBG R135C also has an increased interaction with fibulin-2. Two different substitutions within the dimer interface at R123 (R123H and R123C) reduce the affinity for 5α-dihydrotestosterone, while increasing the relative binding affinity for estradiol. SHBG T48I is defective in calcium binding, which leads to a defect in dimerization, reduced affinity for sex steroids, and an enhanced interaction with fibulin-2, which can all be restored by calcium supplementation. These naturally occurring mutants provide insight into SHBG structure and function, and defects in SHBG production or function need to be considered in the context of its utility as a biomarker of diseases.
Figures
Comment in
-
Reproductive endocrinology: functional effects of sex hormone-binding globulin variants.Nat Rev Endocrinol. 2014 Sep;10(9):516-7. doi: 10.1038/nrendo.2014.120. Epub 2014 Jul 22. Nat Rev Endocrinol. 2014. PMID: 25048036
References
-
- Hammond GL. Access of reproductive steroids to target tissues. Obstet Gynecol Clin North Am. 2002;29:411–423. - PubMed
-
- Hammond GL, Wu TS, Simard M. Evolving utility of sex hormone-binding globulin measurements in clinical medicine. Curr Opin Endocrinol Diabetes Obes. 2012;19:183–189. - PubMed
-
- Hammond GL, Underhill DA, Smith CL, et al. The cDNA-deduced primary structure of human sex hormone-binding globulin and location of its steroid-binding domain. FEBS Lett. 1987;215:100–104. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
