

Centrin3 in trypanosomes maintains the stability of a flagellar inner-arm dynein for cell motility
- PMID: 24892844
- PMCID: PMC4076704
- DOI: 10.1038/ncomms5060
Centrin3 in trypanosomes maintains the stability of a flagellar inner-arm dynein for cell motility
Abstract
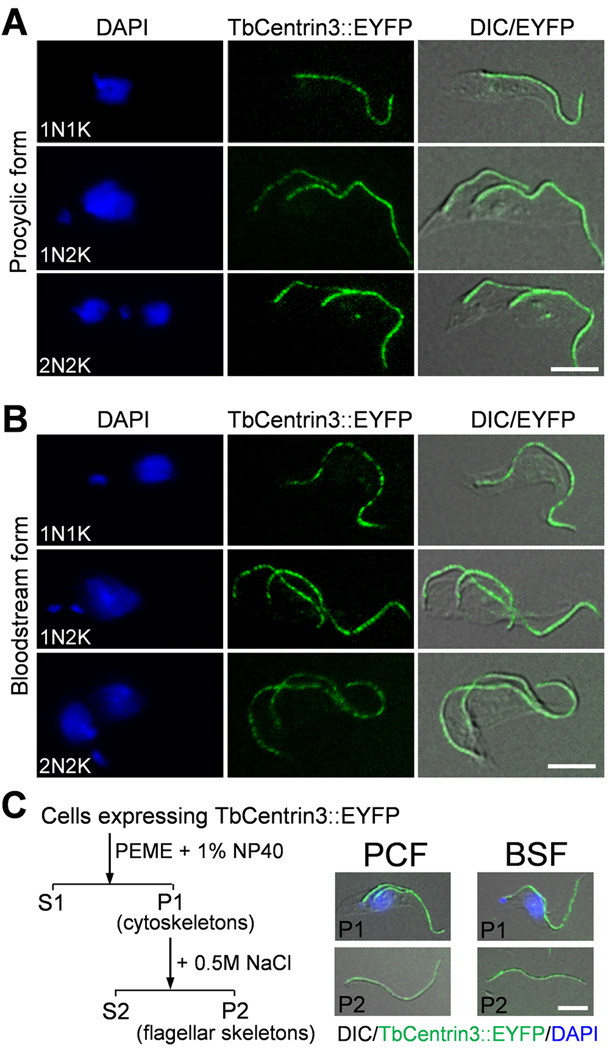
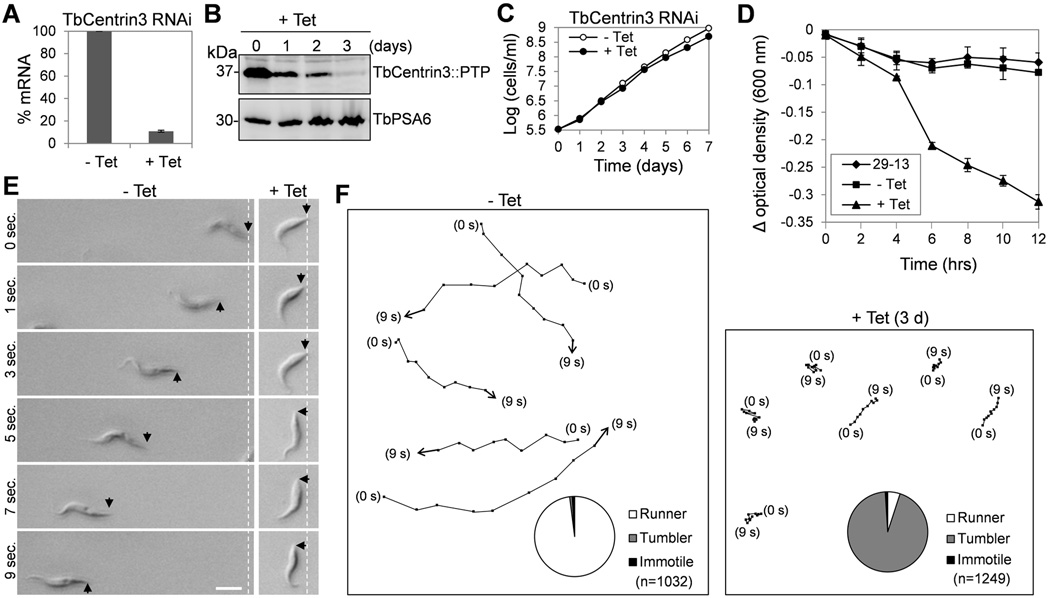
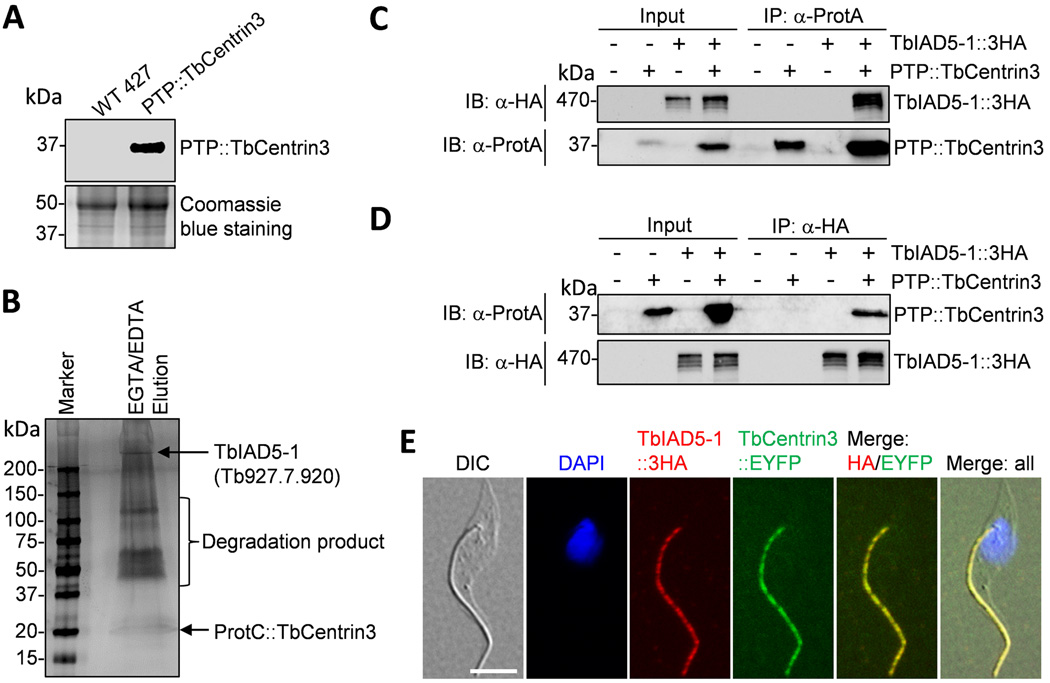
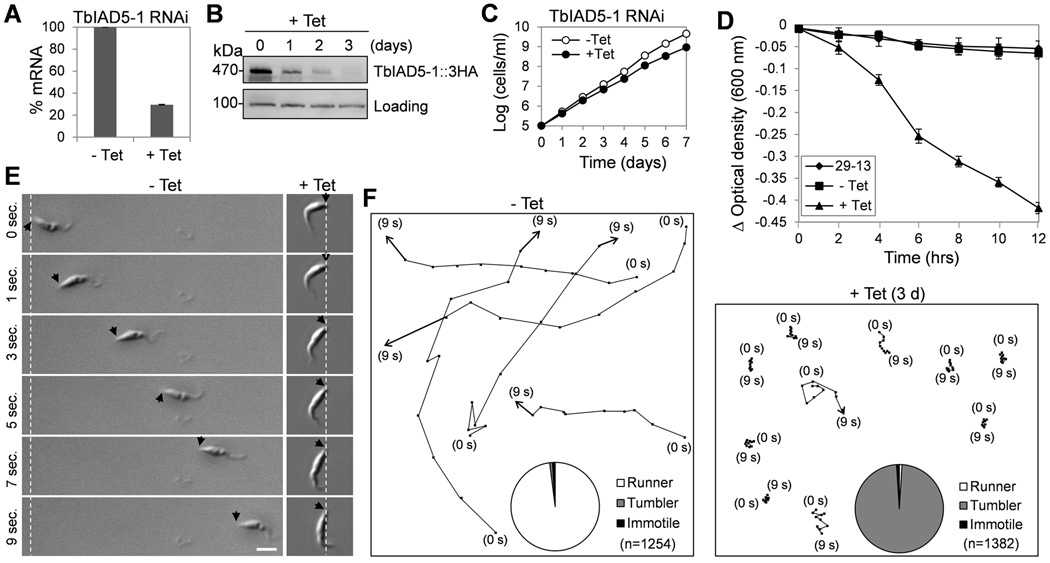
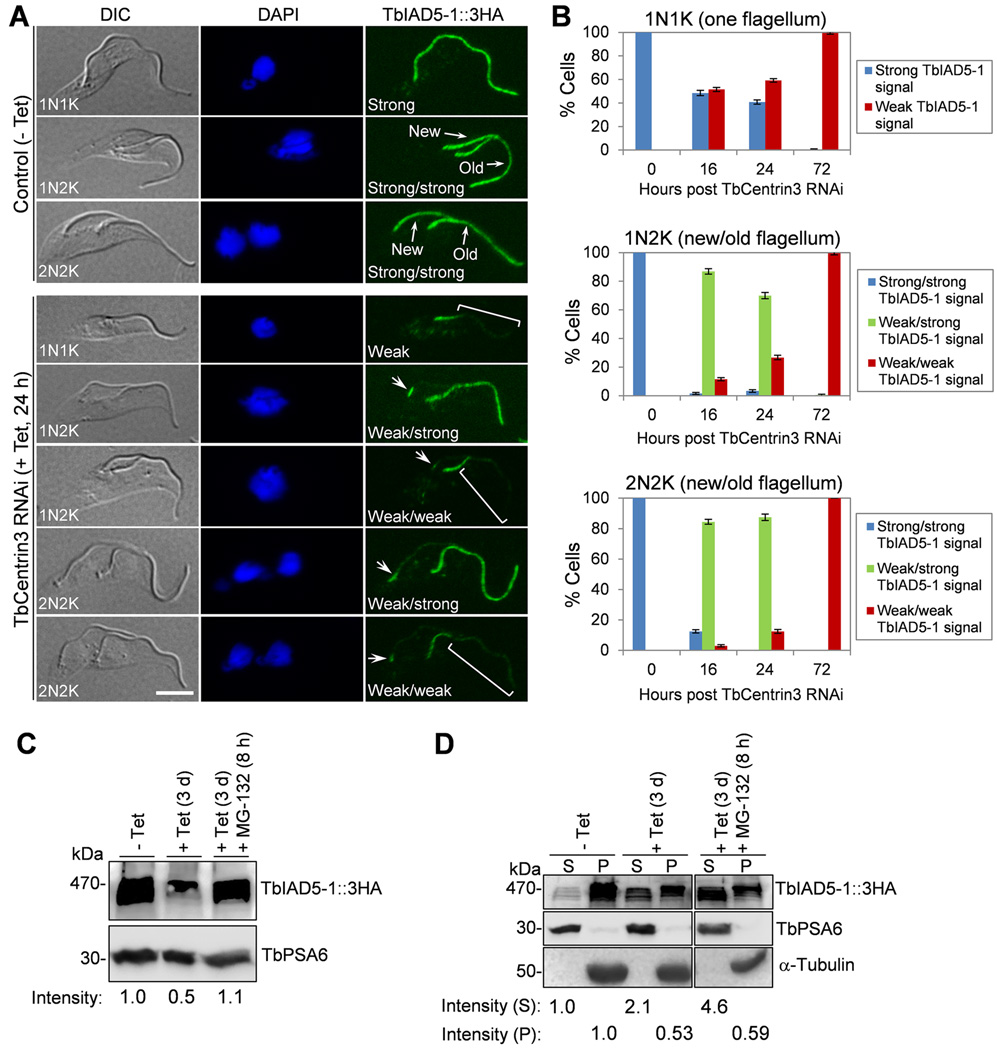
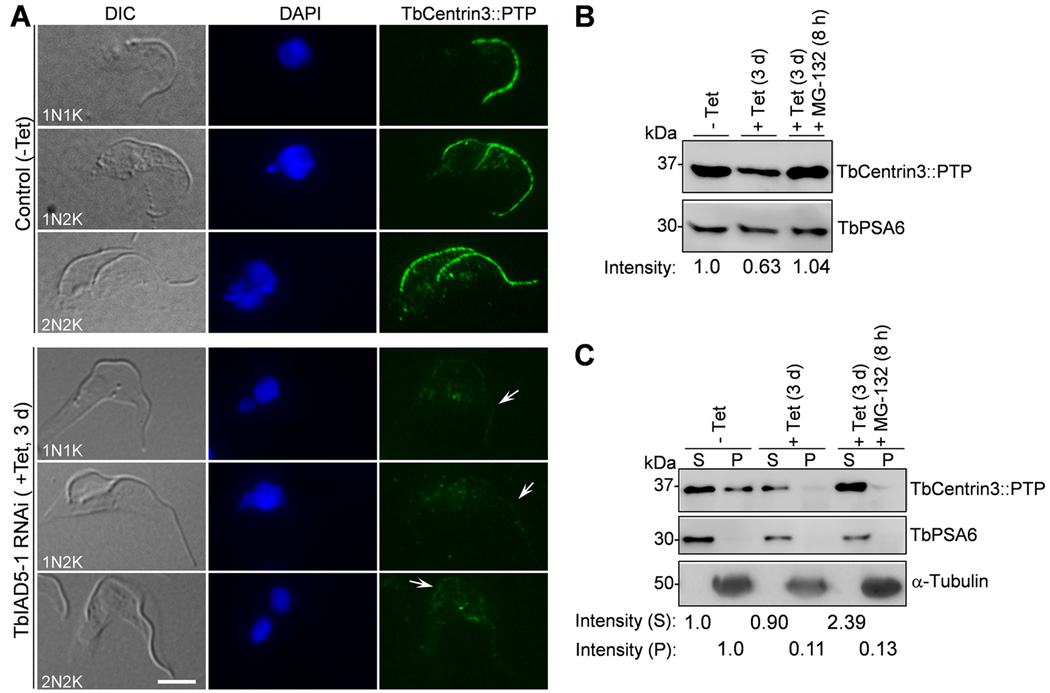
Centrin is a conserved component of centrioles in animals and basal bodies in flagellated organisms. It also associates with axonemal inner-arm dyneins and regulates cell motility, but the underlying mechanism remains elusive. In Trypanosoma brucei, three of the five centrins associate with the flagellar basal body, but no centrin has been found to regulate flagellar motility. Here we show that TbCentrin3 is a flagellar protein and knockdown of TbCentrin3 compromises cell motility. Tandem affinity purification followed by mass spectrometry identifies an inner-arm dynein, TbIAD5-1, as the TbCentrin3 partner, and knockdown of TbIAD5-1 causes similar cell motility defect. Further, we demonstrate the interdependence of TbCentrin3 and TbIAD5-1 for maintaining a stable complex in the flagellar axoneme. Together, these results identify the essential role of TbCentrin3 in cell motility by maintaining the stability of an inner-arm dynein in the flagellum, which may be shared by all the centrin-containing flagellated and ciliated organisms.
Figures
References
-
- Koblenz B, Schoppmeier J, Grunow A, Lechtreck KF. Centrin deficiency in Chlamydomonas causes defects in basal body replication, segregation and maturation. J Cell Sci. 2003;116:2635–2646. - PubMed
-
- Melkonian M. Centrin-mediated cell motility: a novel cell motility mechanism in eukaryotic cells. Bot Acta. 1989;102:3–4.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
