

Cryptosporidium parvum has an active hypusine biosynthesis pathway
- PMID: 24893338
- PMCID: PMC4176827
- DOI: 10.1016/j.molbiopara.2014.05.005
Cryptosporidium parvum has an active hypusine biosynthesis pathway
Abstract
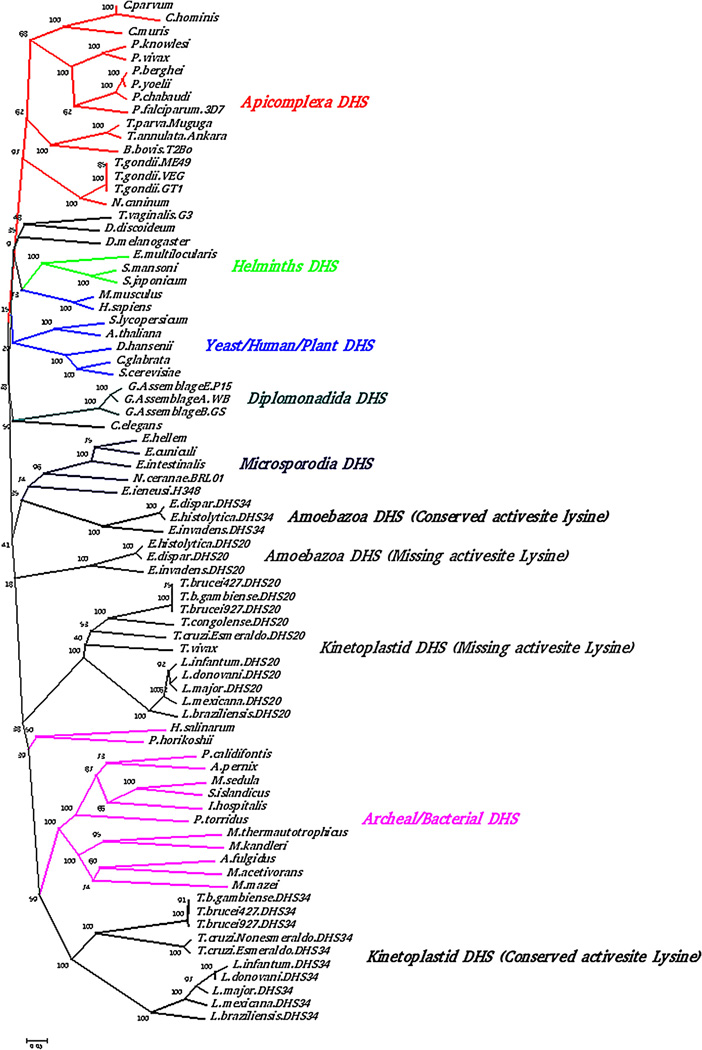
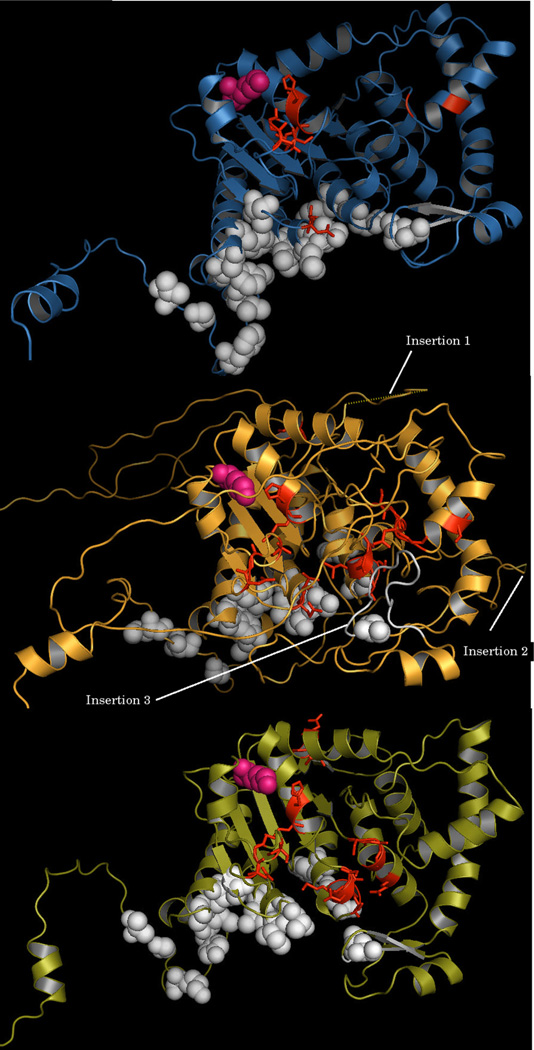
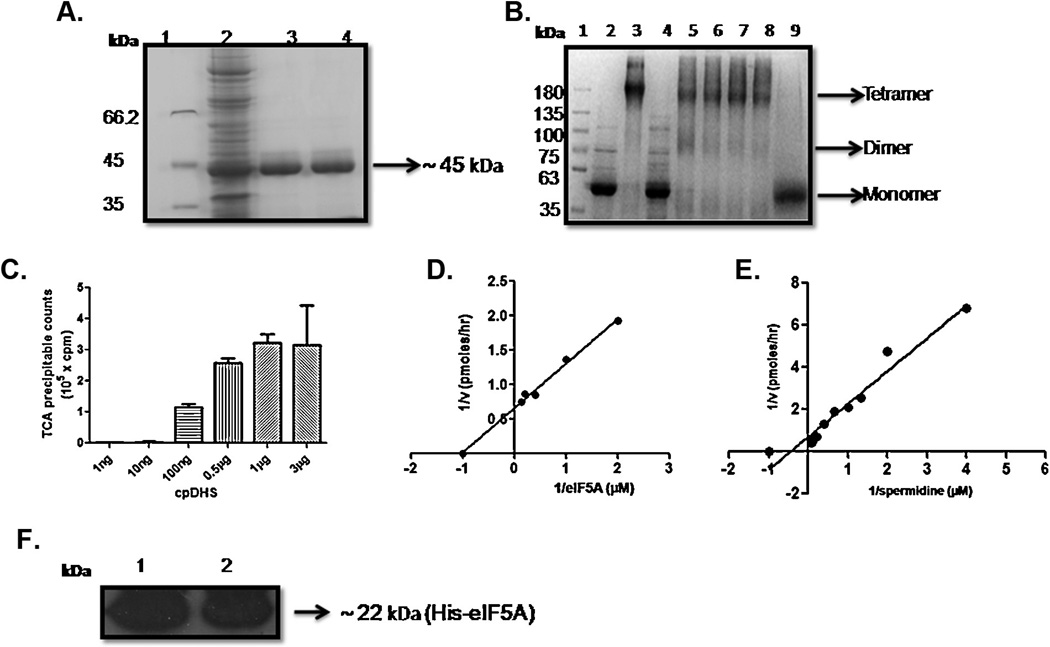
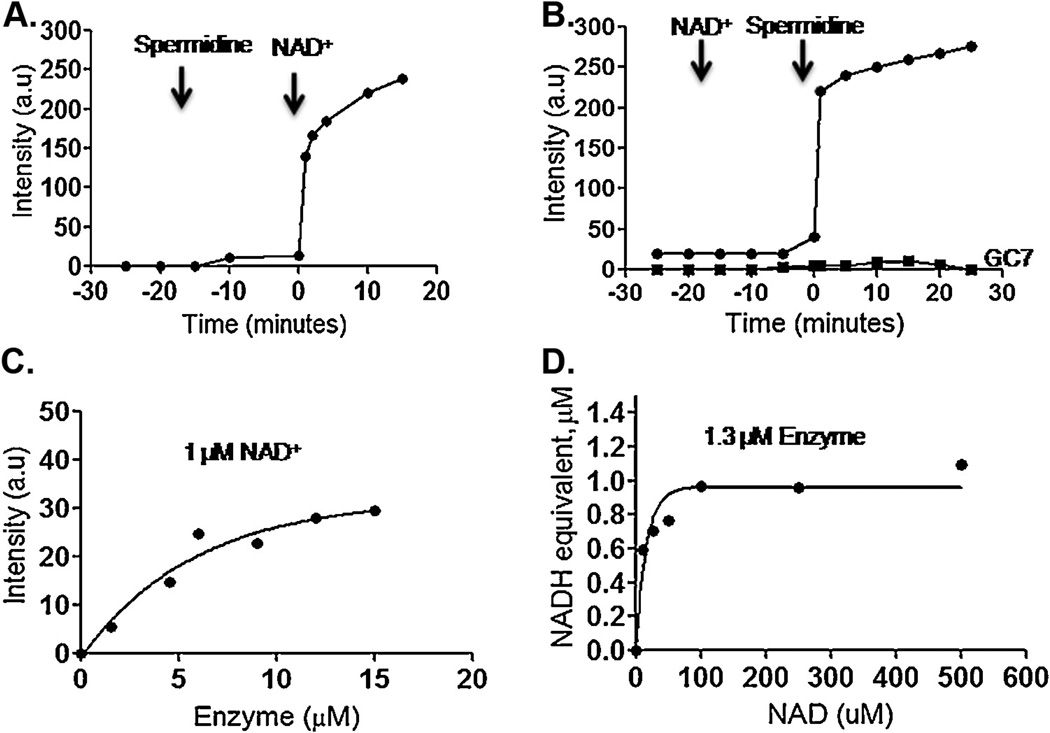
The protozoan parasite Cryptosporidium parvum causes severe enteric infection and diarrheal disease with substantial morbidity and mortality in untreated AIDS patients and children in developing or resource-limited countries. No fully effective treatment is available. Hypusination of eIF5A is an important post-translational modification essential for cell proliferation. This modification occurs in a two step process catalyzed by deoxyhypusine synthase (DHS) followed by deoxyhypusine hydroxylase. An ORF of 1086bp was identified in the C. parvum (Cp) genome which encodes for a putative polypeptide of 362 amino acids. The recombinant CpDHS protein was purified to homogeneity and used to probe the enzyme's mechanism, structure, and inhibition profile in a series of kinetic experiments. Sequence analysis and structural modeling of CpDHS were performed to probe differences with respect to the DHS of other species. Unlike Leishmania, Trypanosomes and Entamoeba, Cryptosporidium contains only a single gene for DHS. Phylogenetic analysis shows that CpDHS is more closely related to apicomplexan DHS than kinetoplastid DHS. Important residues that are essential for the functioning of the enzyme including NAD(+) binding residues, spermidine binding residues and the active site lysine are conserved between CpDHS and human DHS. N(1)-guanyl-1,7-diaminoheptane (GC7), a potent inhibitor of DHS caused an effective inhibition of infection and growth of C. parvum in HCT-8 cells.
Keywords: Cryptosporidium parvum; Deoxyhypsuine synthase; Hypusine pathway; Protozoan parasite.
Copyright © 2014 Elsevier B.V. All rights reserved.
Figures

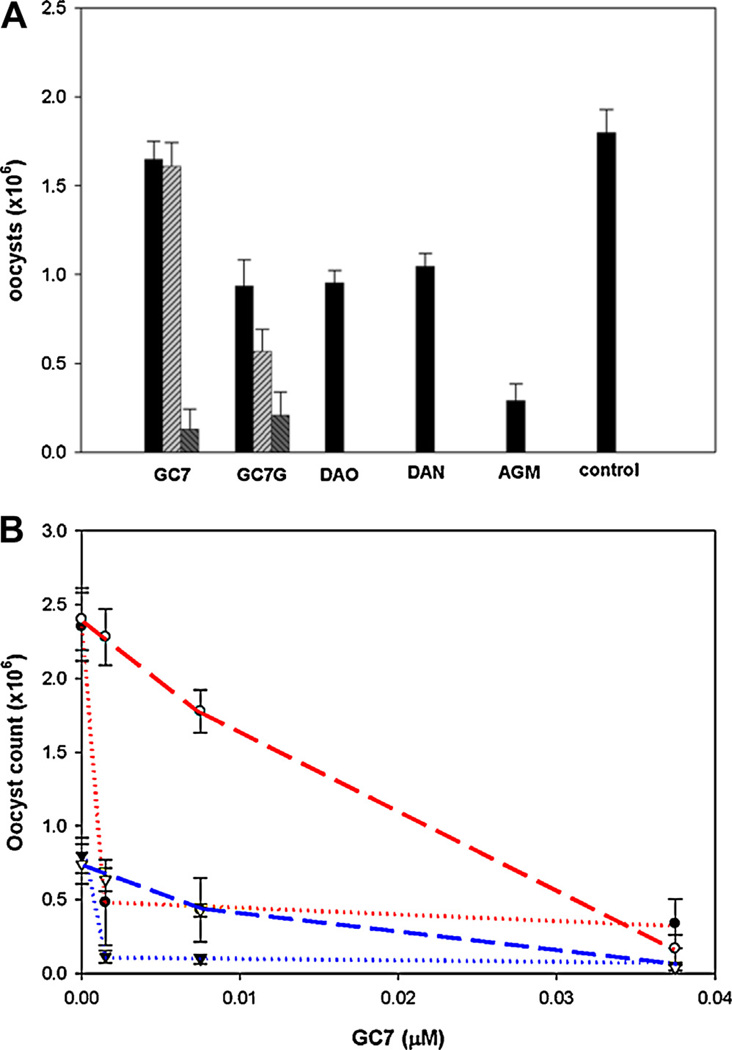
 ), 0.04 µM (
), 0.04 µM ( ), or 0.021 µM (
), or 0.021 µM ( ) GC7; 0.45 µM (), 0.90 µM (), or 4.5 µM () GC7G; 24µM diaminooctane (DAO); 36 µM diaminononane (DAN); 10 µM agmatine (AGM). Results are expressed as + SD of triplicate experiments compared to control oocysts lacking added compounds. Paromomycin (150 µM) caused 58% inhibition of C. parvum oocysts production. (B) Inhibition of C. parvum infection and growth by GC7. C. parvum oocyst production after 24 h incubation with HCT-8 cells. Oocysts (
) GC7; 0.45 µM (), 0.90 µM (), or 4.5 µM () GC7G; 24µM diaminooctane (DAO); 36 µM diaminononane (DAN); 10 µM agmatine (AGM). Results are expressed as + SD of triplicate experiments compared to control oocysts lacking added compounds. Paromomycin (150 µM) caused 58% inhibition of C. parvum oocysts production. (B) Inhibition of C. parvum infection and growth by GC7. C. parvum oocyst production after 24 h incubation with HCT-8 cells. Oocysts ( ) or sporozoites (
) or sporozoites ( ) preincubated for 1.5 h with HCT-8 cells prior to addition of GC7; Oocysts (
) preincubated for 1.5 h with HCT-8 cells prior to addition of GC7; Oocysts ( ) or sporozoites (
) or sporozoites ( ) preincubated with GC7 for 30 min prior to infecting HCT-8 cells.
) preincubated with GC7 for 30 min prior to infecting HCT-8 cells.Similar articles
-
A unique modification of the eukaryotic initiation factor 5A shows the presence of the complete hypusine pathway in Leishmania donovani.PLoS One. 2012;7(3):e33138. doi: 10.1371/journal.pone.0033138. Epub 2012 Mar 16. PLoS One. 2012. PMID: 22438895 Free PMC article.
-
Cloning, expression and functional activity of deoxyhypusine synthase from Plasmodium vivax.BMC Microbiol. 2006 Oct 16;6:91. doi: 10.1186/1471-2180-6-91. BMC Microbiol. 2006. PMID: 17042947 Free PMC article.
-
Immunolocation and enzyme activity analysis of Cryptosporidium parvum enolase.Parasit Vectors. 2017 May 31;10(1):273. doi: 10.1186/s13071-017-2200-y. Parasit Vectors. 2017. PMID: 28569179 Free PMC article.
-
Posttranslational synthesis of hypusine: evolutionary progression and specificity of the hypusine modification.Amino Acids. 2007 Aug;33(2):341-50. doi: 10.1007/s00726-007-0525-0. Epub 2007 May 4. Amino Acids. 2007. PMID: 17476569 Free PMC article. Review.
-
Cryptosporidium: genomic and biochemical features.Exp Parasitol. 2010 Jan;124(1):2-9. doi: 10.1016/j.exppara.2008.12.014. Epub 2008 Dec 31. Exp Parasitol. 2010. PMID: 19187778 Free PMC article. Review.
Cited by
-
Polyamines in protozoan pathogens.J Biol Chem. 2018 Nov 30;293(48):18746-18756. doi: 10.1074/jbc.TM118.003342. Epub 2018 Oct 17. J Biol Chem. 2018. PMID: 30333232 Free PMC article. Review.
-
Structural features and development of an assay platform of the parasite target deoxyhypusine synthase of Brugia malayi and Leishmania major.PLoS Negl Trop Dis. 2020 Oct 12;14(10):e0008762. doi: 10.1371/journal.pntd.0008762. eCollection 2020 Oct. PLoS Negl Trop Dis. 2020. PMID: 33044977 Free PMC article.
References
-
- Fayer R. Cryptosporidium and cryptosporidiosis. CRC Press; 1997.
-
- Semenza JC, Nichols G. Cryptosporidiosis surveillance and water-borne outbreaks in Europe. Euro Surveill. 2007;12:E13–E14. - PubMed
-
- Mead JR. Cryptosporidiosis and the challenges of chemotherapy. Drug Resist Update. 2002;5:47–57. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
