

Vertex models of epithelial morphogenesis
- PMID: 24896108
- PMCID: PMC4052277
- DOI: 10.1016/j.bpj.2013.11.4498
Vertex models of epithelial morphogenesis
Abstract
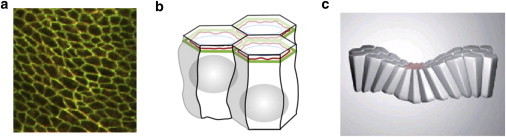
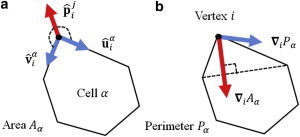
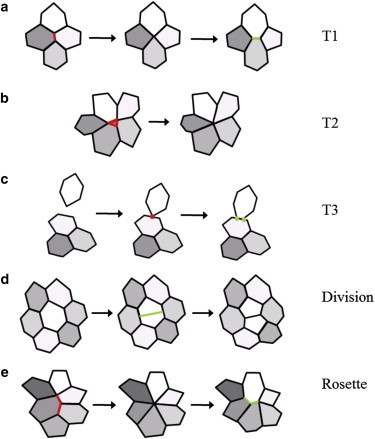
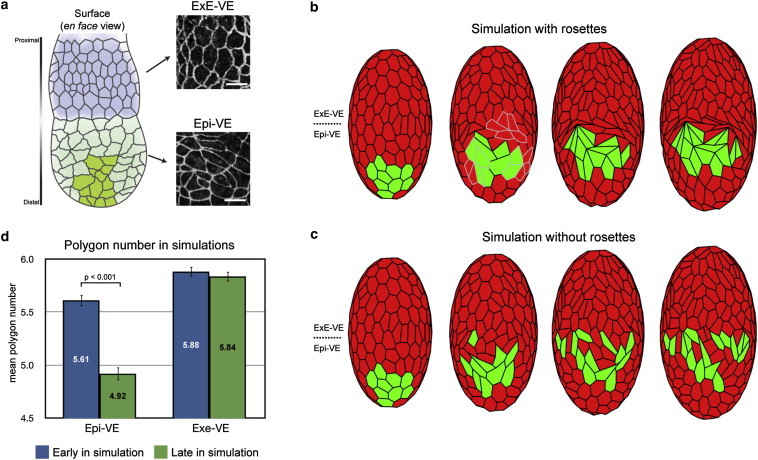
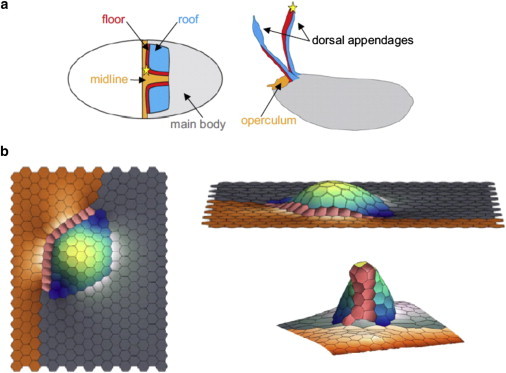
The dynamic behavior of epithelial cell sheets plays a central role during numerous developmental processes. Genetic and imaging studies of epithelial morphogenesis in a wide range of organisms have led to increasingly detailed mechanisms of cell sheet dynamics. Computational models offer a useful means by which to investigate and test these mechanisms, and have played a key role in the study of cell-cell interactions. A variety of modeling approaches can be used to simulate the balance of forces within an epithelial sheet. Vertex models are a class of such models that consider cells as individual objects, approximated by two-dimensional polygons representing cellular interfaces, in which each vertex moves in response to forces due to growth, interfacial tension, and pressure within each cell. Vertex models are used to study cellular processes within epithelia, including cell motility, adhesion, mitosis, and delamination. This review summarizes how vertex models have been used to provide insight into developmental processes and highlights current challenges in this area, including progressing these models from two to three dimensions and developing new tools for model validation.
Copyright © 2014 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Lecuit T., Lenne P.F. Cell surface mechanics and the control of cell shape, tissue patterns and morphogenesis. Nat. Rev. Mol. Cell Biol. 2007;8:633–644. - PubMed
-
- Wallingford J.B. Neural tube closure and neural tube defects: studies in animal models reveal known knowns and known unknowns. Am. J. Med. Genet. C. Semin. Med. Genet. 2005;135C:59–68. - PubMed
-
- Lubarsky B., Krasnow M.A. Tube morphogenesis: making and shaping biological tubes. Cell. 2003;112:19–28. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
