

Development of the embryonic and larval peripheral nervous system of Drosophila
- PMID: 24896657
- PMCID: PMC4318504
- DOI: 10.1002/wdev.135
Development of the embryonic and larval peripheral nervous system of Drosophila
Abstract
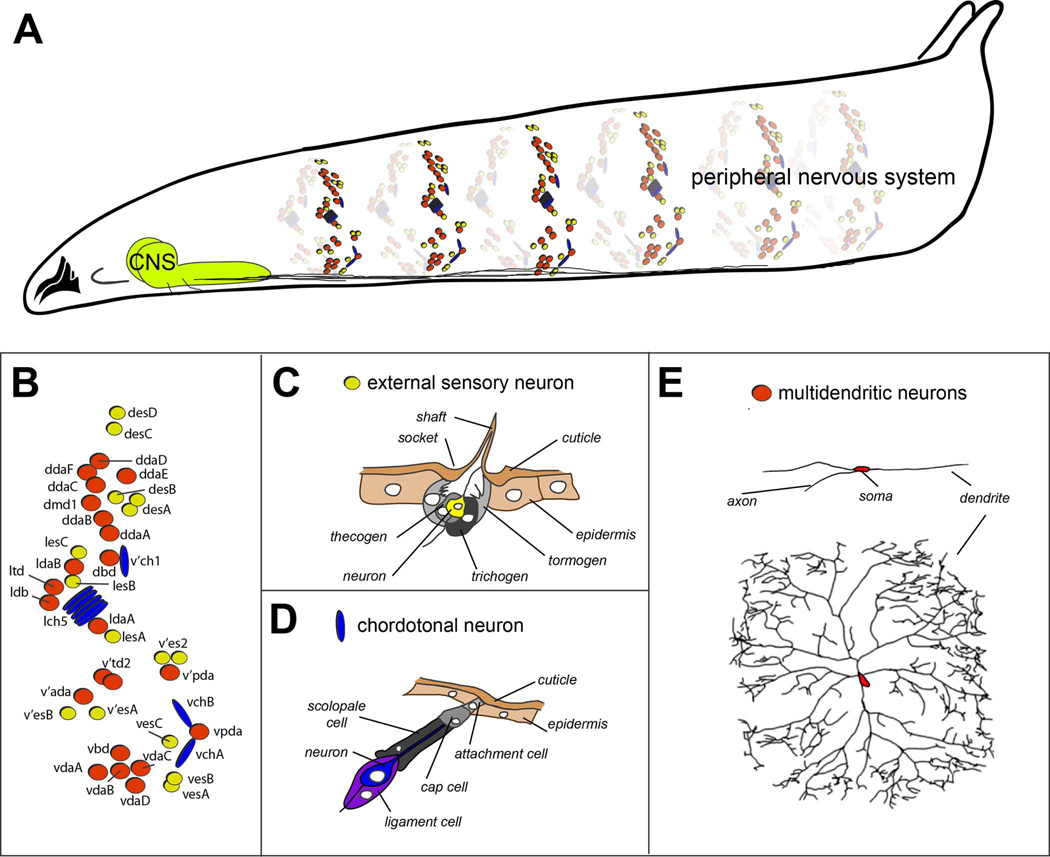
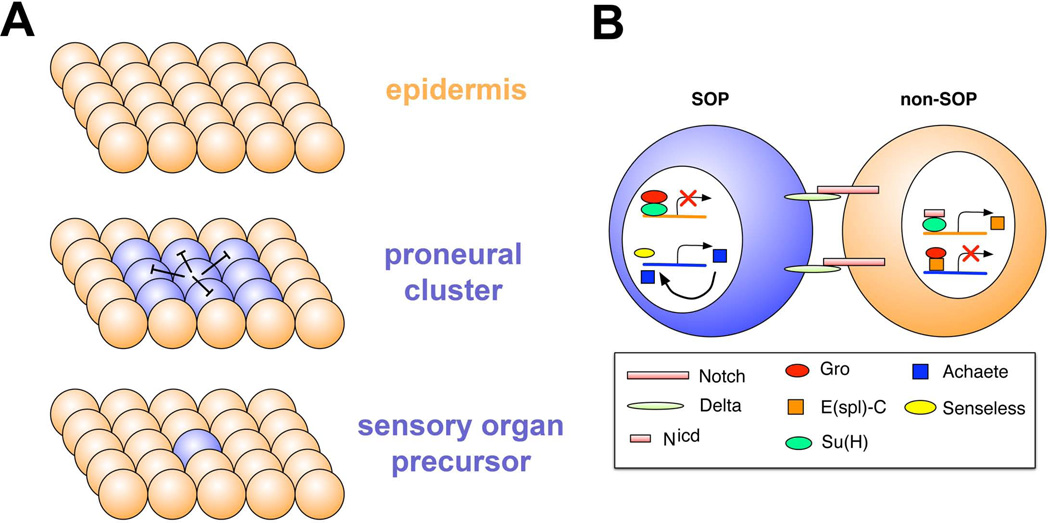
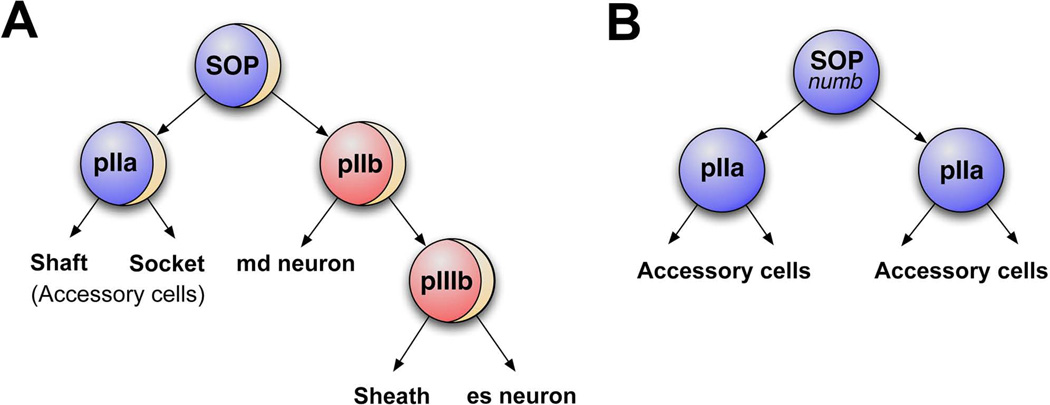
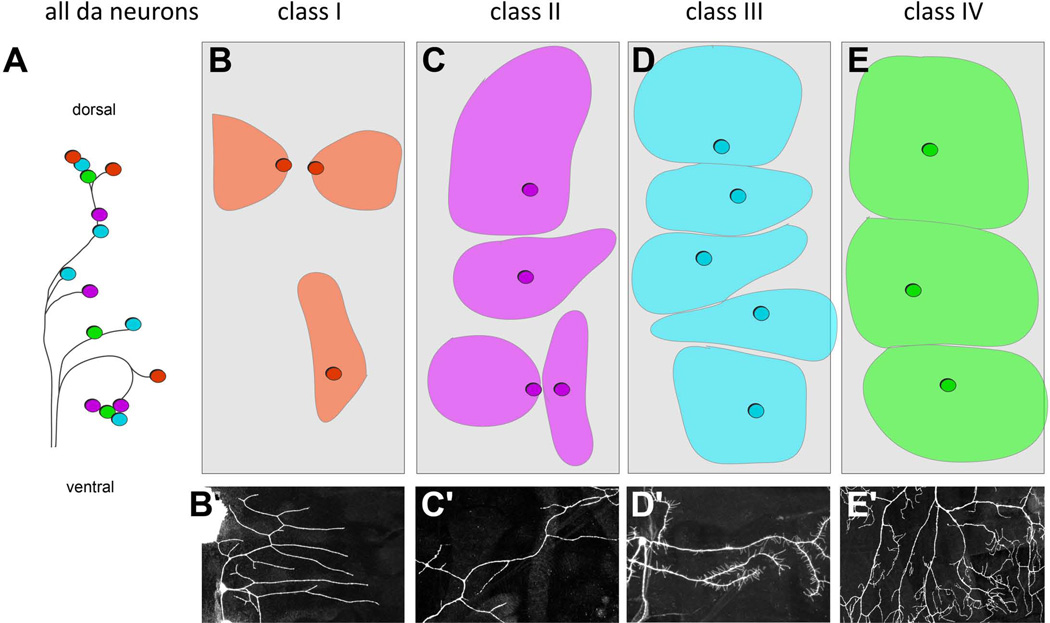
The peripheral nervous system (PNS) of embryonic and larval stage Drosophila consists of diverse types of sensory neurons positioned along the body wall. Sensory neurons, and associated end organs, show highly stereotyped locations and morphologies. Many powerful genetic tools for gene manipulation available in Drosophila make the PNS an advantageous system for elucidating basic principles of neural development. Studies of the Drosophila PNS have provided key insights into molecular mechanisms of cell fate specification, asymmetric cell division, and dendritic morphogenesis. A canonical lineage gives rise to sensory neurons and associated organs, and cells within this lineage are diversified through asymmetric cell divisions. Newly specified sensory neurons develop specific dendritic patterns, which are controlled by numerous factors including transcriptional regulators, interactions with neighboring neurons, and intracellular trafficking systems. In addition, sensory axons show modality specific terminations in the central nervous system, which are patterned by secreted ligands and their receptors expressed by sensory axons. Modality-specific axon projections are critical for coordinated larval behaviors. We review the molecular basis for PNS development and address some of the instances in which the mechanisms and molecules identified are conserved in vertebrate development.
© 2014 Wiley Periodicals, Inc.
Figures
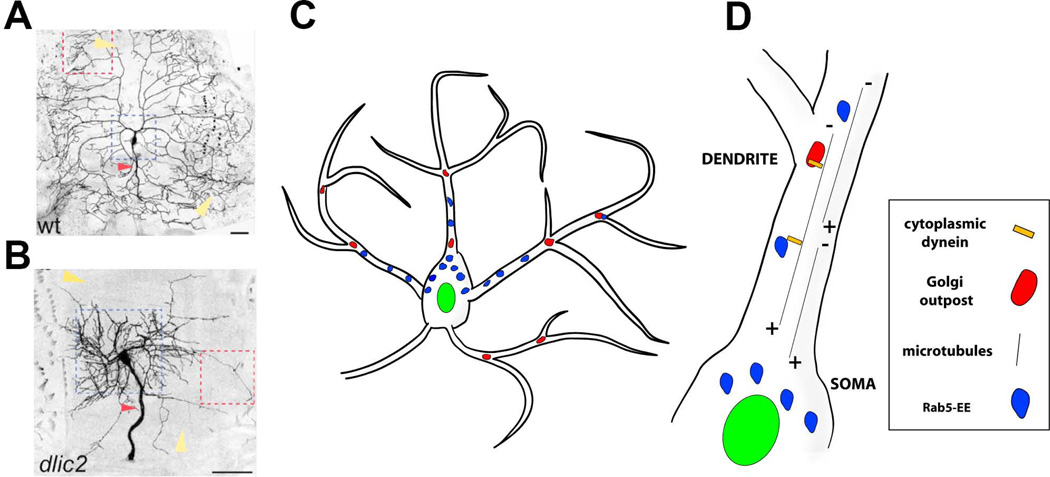
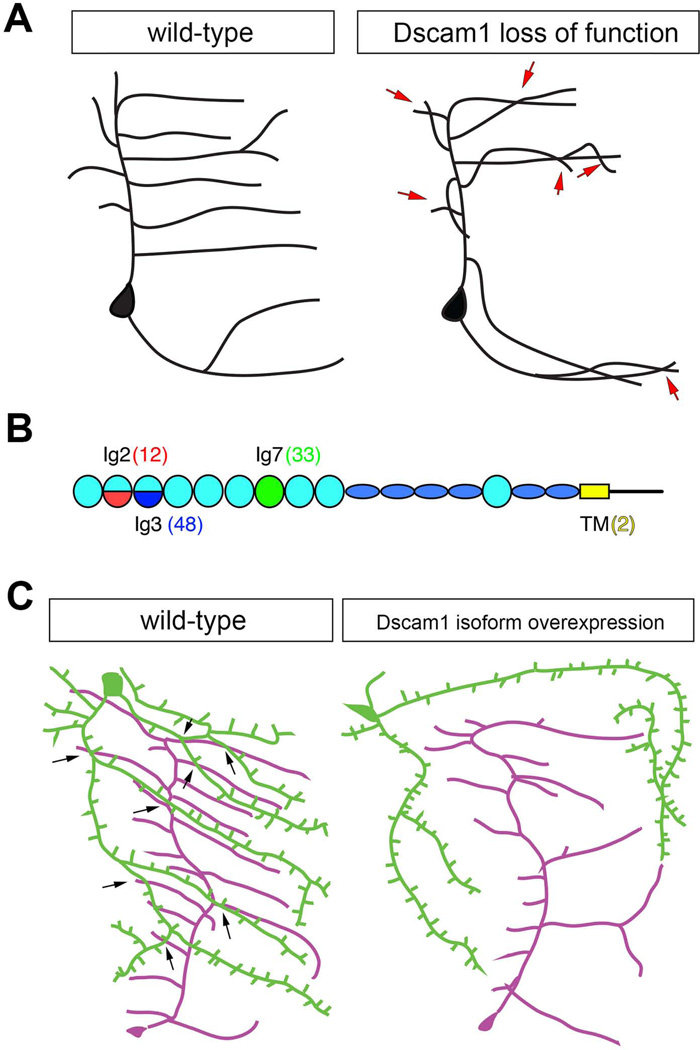
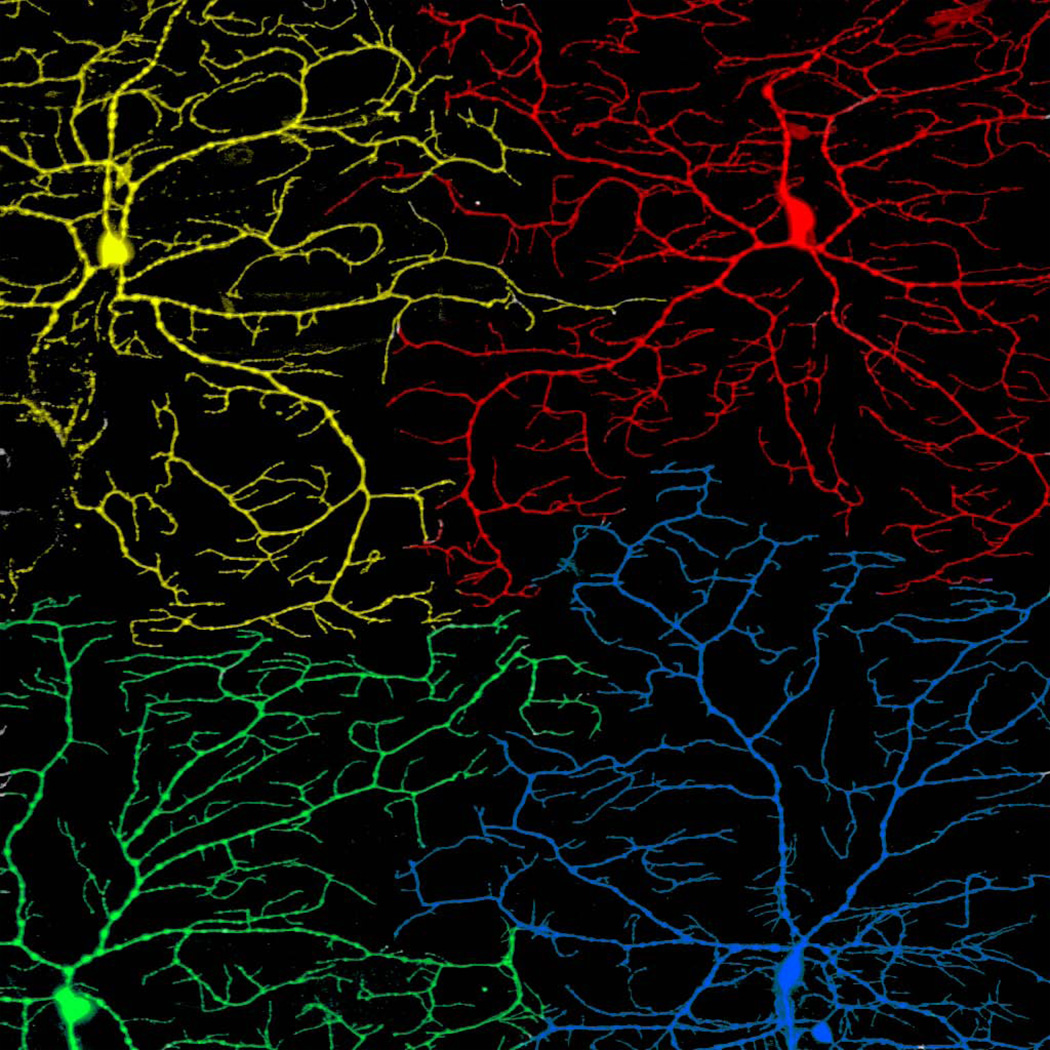
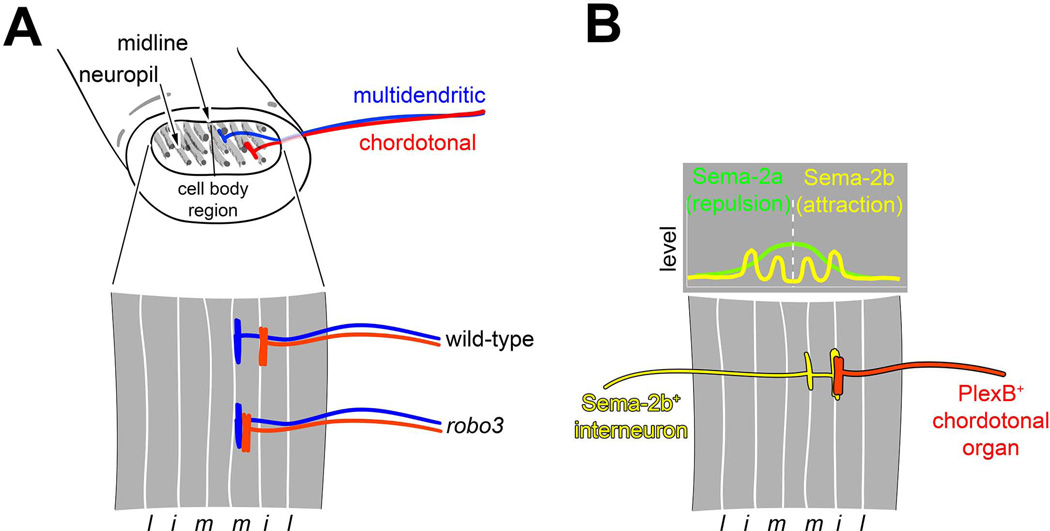
Similar articles
-
Lineage, cell polarity and inscuteable function in the peripheral nervous system of the Drosophila embryo.Development. 2001 Mar;128(5):631-43. doi: 10.1242/dev.128.5.631. Development. 2001. PMID: 11171389
-
Projections of Drosophila multidendritic neurons in the central nervous system: links with peripheral dendrite morphology.Development. 2007 Jan;134(1):55-64. doi: 10.1242/dev.02666. Development. 2007. PMID: 17164414
-
Central projections of sensory neurons in the Drosophila embryo correlate with sensory modality, soma position, and proneural gene function.J Neurosci. 1995 Mar;15(3 Pt 1):1755-67. doi: 10.1523/JNEUROSCI.15-03-01755.1995. J Neurosci. 1995. PMID: 7891133 Free PMC article.
-
Evolutionarily conserved mechanisms regulating neural development: lessons from the development of Drosophila peripheral nervous systems.Hum Cell. 1997 Sep;10(3):139-50. Hum Cell. 1997. PMID: 9436033 Review.
-
Morphogenesis of Drosophila melanogaster macrochaetes: cell fate determination for bristle organ.J Stem Cells. 2012;7(1):19-41. J Stem Cells. 2012. PMID: 23550342 Review.
Cited by
-
Macrophages and Their Organ Locations Shape Each Other in Development and Homeostasis - A Drosophila Perspective.Front Cell Dev Biol. 2021 Mar 11;9:630272. doi: 10.3389/fcell.2021.630272. eCollection 2021. Front Cell Dev Biol. 2021. PMID: 33777939 Free PMC article. Review.
-
Expression of a human variant of CHMP2B linked to neurodegeneration in Drosophila external sensory organs leads to cell fate transformations associated with increased Notch activity.Dev Neurobiol. 2020 Mar;80(3-4):85-97. doi: 10.1002/dneu.22722. Epub 2019 Oct 23. Dev Neurobiol. 2020. PMID: 31587468 Free PMC article.
-
Dynamic instability of dendrite tips generates the highly branched morphologies of sensory neurons.Sci Adv. 2022 Jul;8(26):eabn0080. doi: 10.1126/sciadv.abn0080. Epub 2022 Jun 29. Sci Adv. 2022. PMID: 35767611 Free PMC article.
-
Neural circuits driving larval locomotion in Drosophila.Neural Dev. 2018 Apr 19;13(1):6. doi: 10.1186/s13064-018-0103-z. Neural Dev. 2018. PMID: 29673388 Free PMC article. Review.
-
Even-Skipped(+) Interneurons Are Core Components of a Sensorimotor Circuit that Maintains Left-Right Symmetric Muscle Contraction Amplitude.Neuron. 2015 Oct 21;88(2):314-29. doi: 10.1016/j.neuron.2015.09.009. Epub 2015 Oct 1. Neuron. 2015. PMID: 26439528 Free PMC article.
References
-
- McIver SB. Mechanoreception. In: Kerkut GA, Gilbert LI, editors. Comprehensive Insect Physiology Biochemistry and Pharmacology. Vol. 6. Oxford: Elsevier; 1985. pp. 71–132.
-
- Hartenstein V. Development of Insect Sensilla. In: Gilbert LI, Iatrou K, Gill SS, editors. Comprehensive Molecular Insect Science. Vol. 1. Elsevier; 2005. pp. 379–419.
-
- Bodmer R, Jan YN. Morphological differentiation of the embryonic peripheral neurons in Drosophila. Roux's Arch Dev Biol. 1987;196:69–77. - PubMed
-
- Grueber WB, Jan LY, Jan YN. Tiling of the Drosophila epidermis by multidendritic sensory neurons. Development. 2002;129:2867–2878. - PubMed
-
- Sweeney NT, Li W, Gao FB. Genetic manipulation of single neurons in vivo reveals specific roles of flamingo in neuronal morphogenesis. Dev Biol. 2002;247:76–88. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
