

Stress-induced start codon fidelity regulates arsenite-inducible regulatory particle-associated protein (AIRAP) translation
- PMID: 24898249
- PMCID: PMC4110281
- DOI: 10.1074/jbc.M114.547828
Stress-induced start codon fidelity regulates arsenite-inducible regulatory particle-associated protein (AIRAP) translation
Abstract
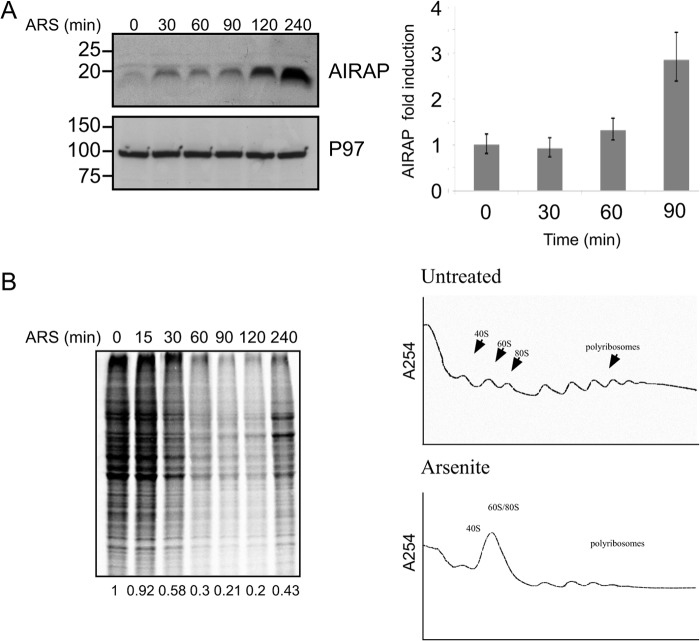
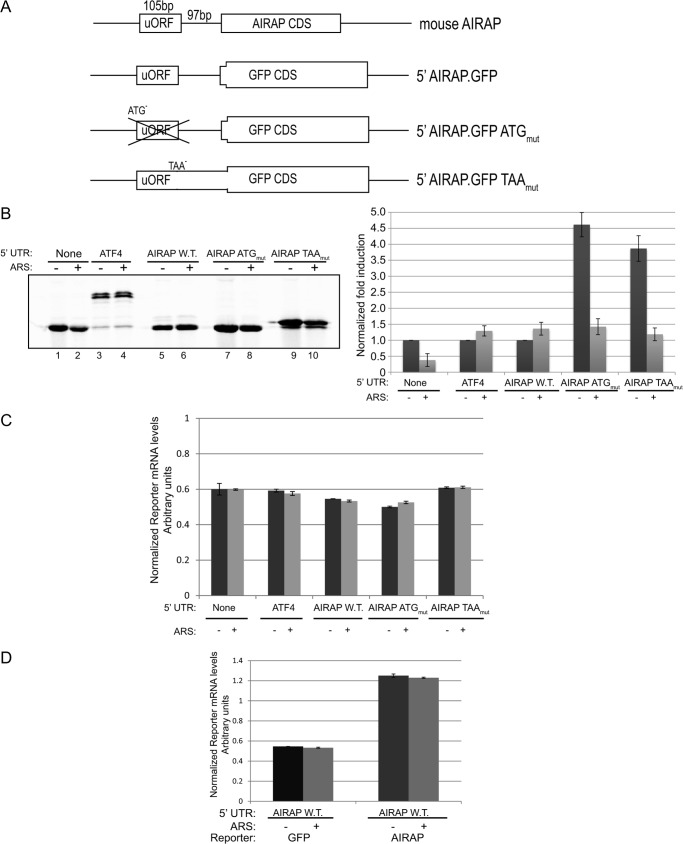
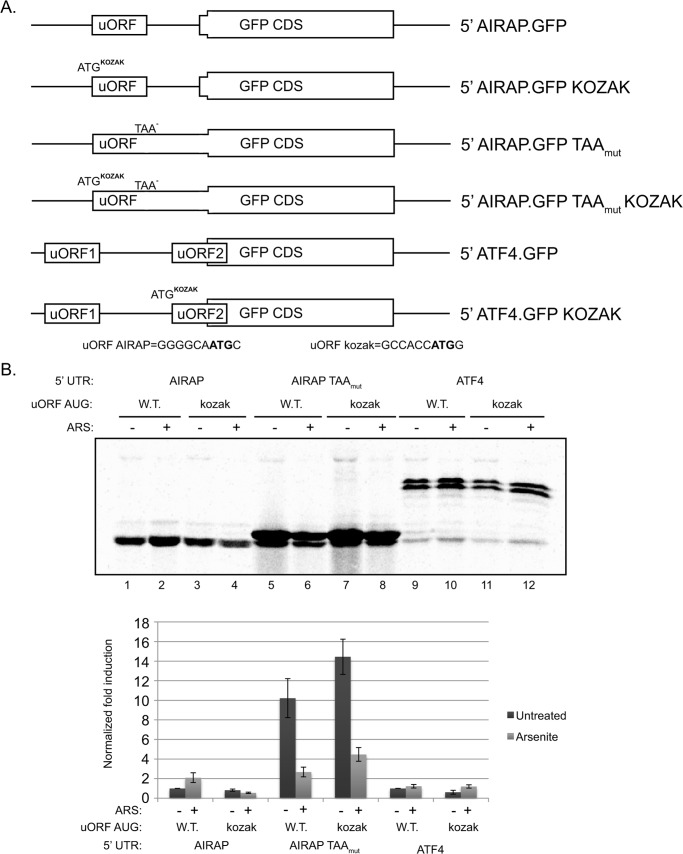
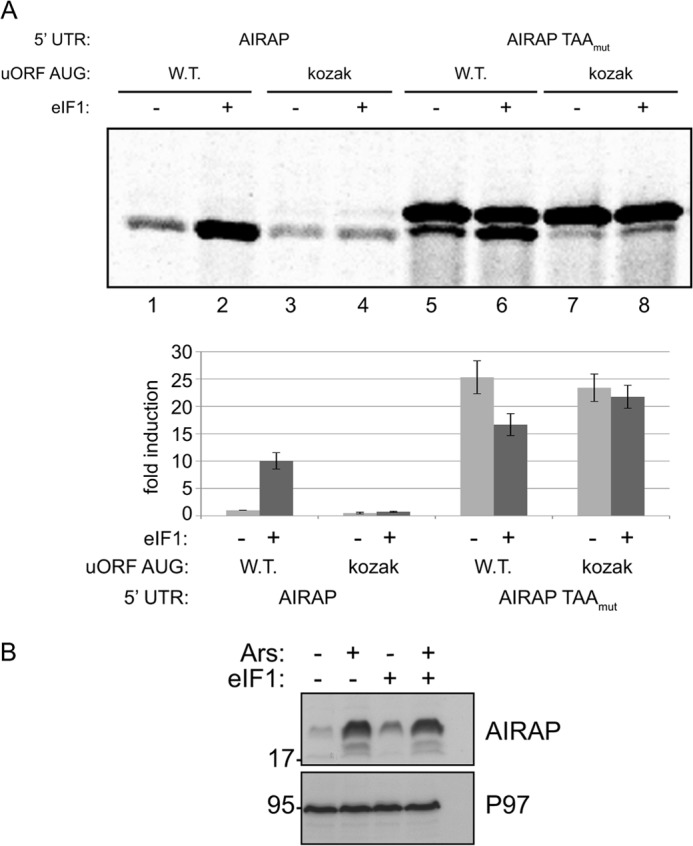
Initial steps in protein synthesis are highly regulated processes as they define the reading frame of the translation machinery. Eukaryotic translation initiation is a process facilitated by numerous factors (eIFs), aimed to form a "scanning" mechanism toward the initiation codon. Translation initiation of the main open reading frame (ORF) in an mRNA transcript has been reported to be regulated by upstream open reading frames (uORFs) in a manner of re-initiation. This mode of regulation is governed by the phosphorylation status of eIF2α and controlled by cellular stresses. Another mode of translational initiation regulation is leaky scanning, and this regulatory process has not been extensively studied. We have identified arsenite- inducible regulatory particle-associated protein (AIRAP) transcript to be translationally induced during arsenite stress conditions. AIRAP transcript contains a single uORF in a poor-kozak context. AIRAP translation induction is governed by means of leaky scanning and not re-initiation. This induction of AIRAP is solely dependent on eIF1 and the uORF kozak context. We show that eIF1 is phosphorylated under specific conditions that induce protein misfolding and have biochemically characterized this site of phosphorylation. Our data indicate that leaky scanning like re-initiation is responsive to stress conditions and that leaky scanning can induce ORF translation by bypassing poor kozak context of a single uORF transcript.
Figures
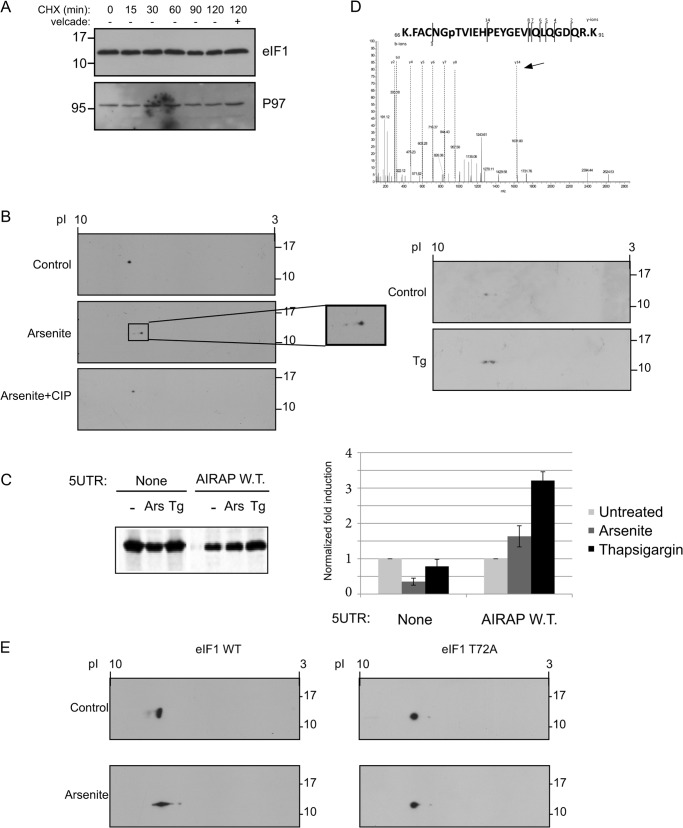
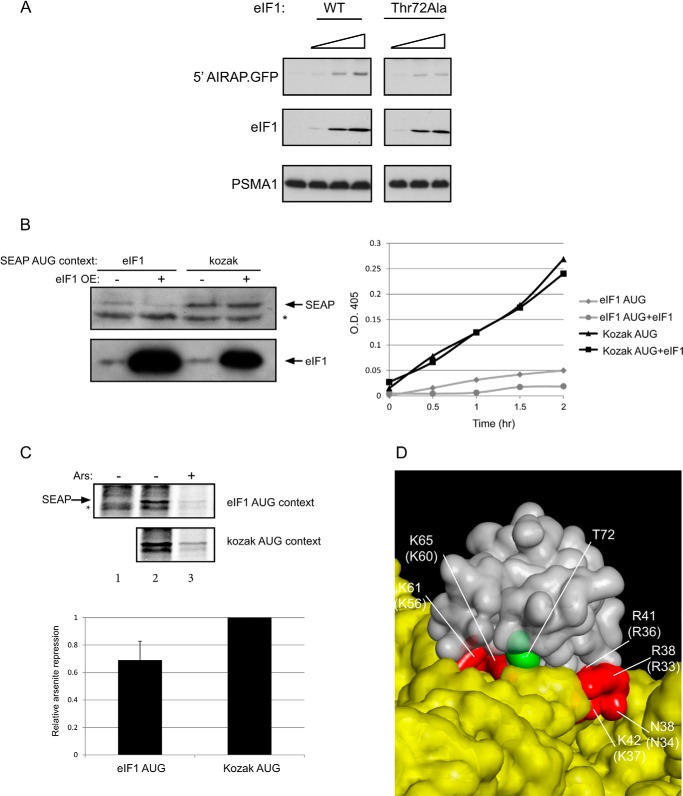
References
-
- Chaudhuri J., Chowdhury D., Maitra U. (1999) Distinct functions of eukaryotic translation initiation factors eIF1A and eIF3 in the formation of the 40 S ribosomal preinitiation complex. J. Biol. Chem. 274, 17975–17980 - PubMed
-
- Kozak M. (1991) Structural features in eukaryotic mRNAs that modulate the initiation of translation. J. Biol. Chem. 266, 19867–19870 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
