

Chronic opioids regulate KATP channel subunit Kir6.2 and carbonic anhydrase I and II expression in rat adrenal chromaffin cells via HIF-2α and protein kinase A
- PMID: 24898587
- PMCID: PMC4121586
- DOI: 10.1152/ajpcell.00135.2014
Chronic opioids regulate KATP channel subunit Kir6.2 and carbonic anhydrase I and II expression in rat adrenal chromaffin cells via HIF-2α and protein kinase A
Abstract
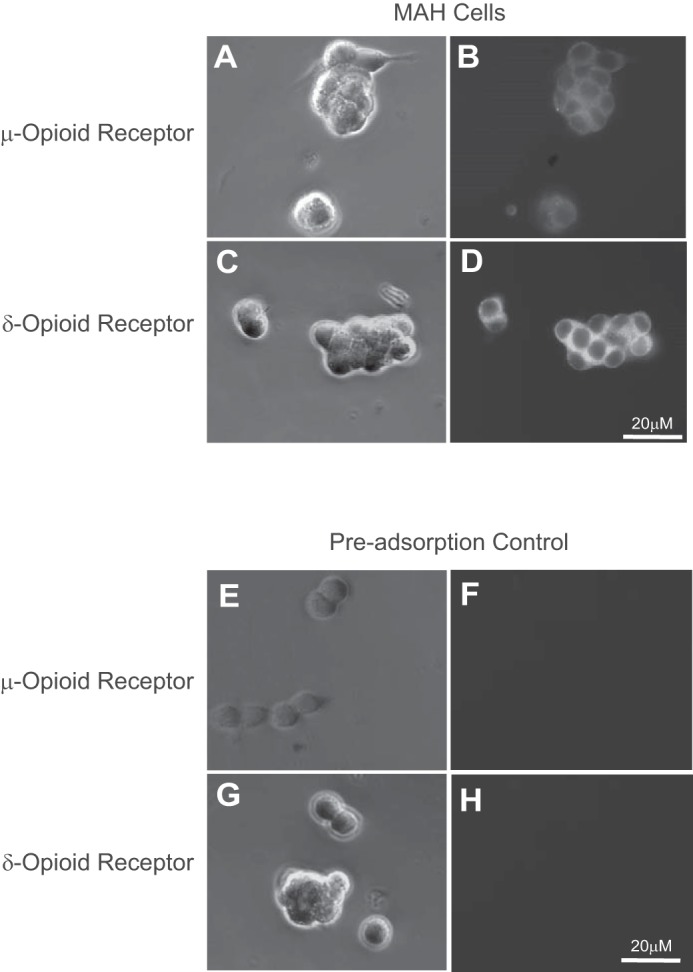
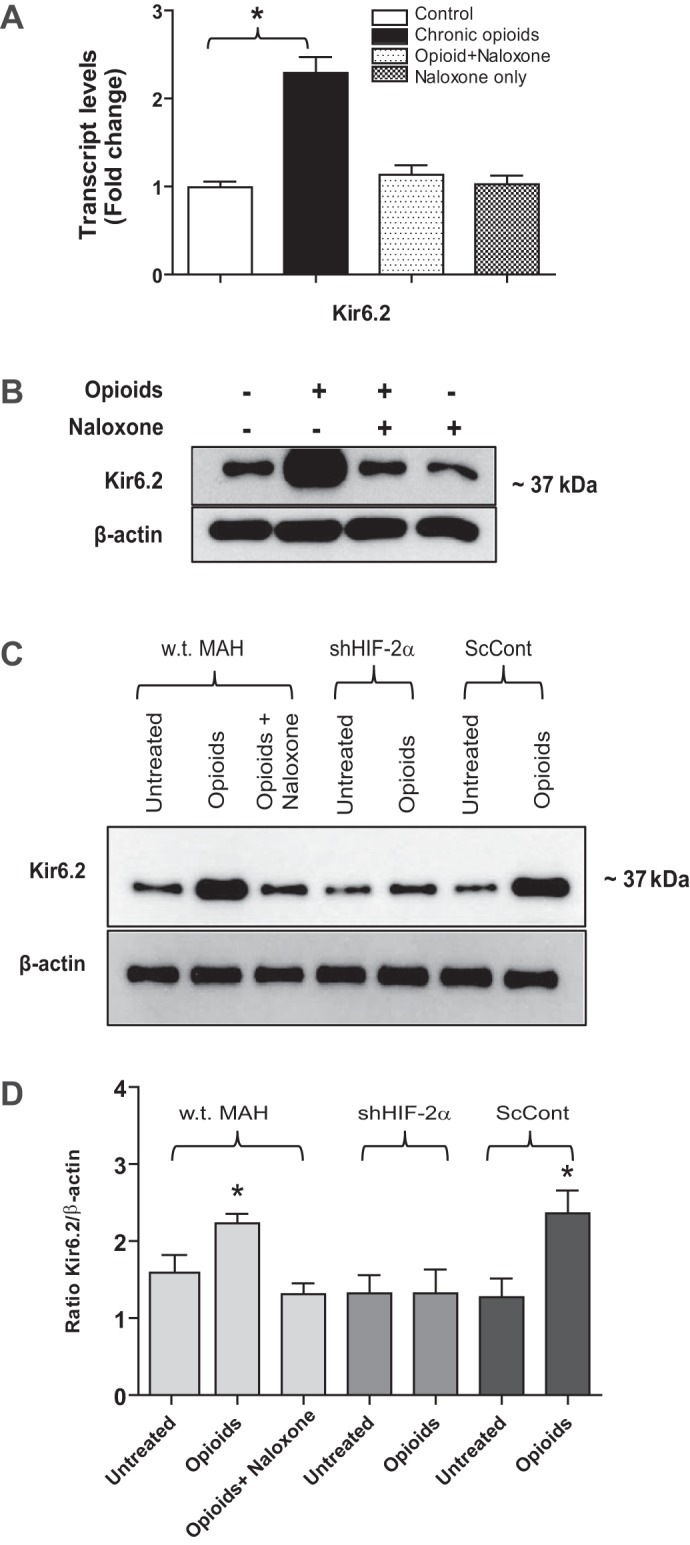
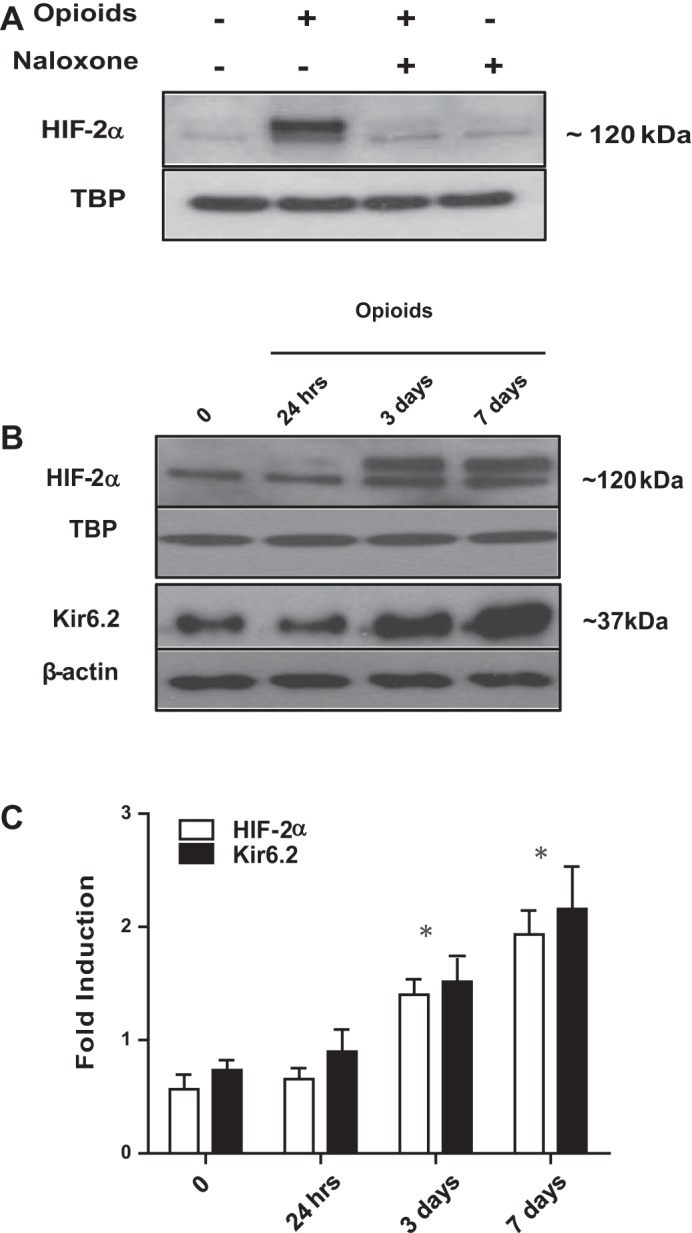
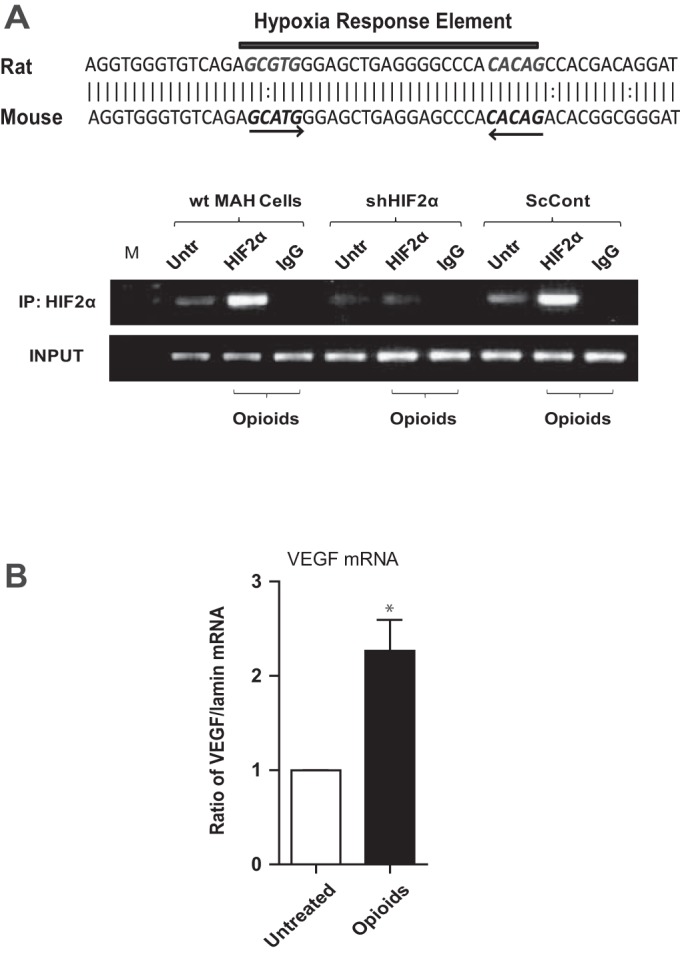
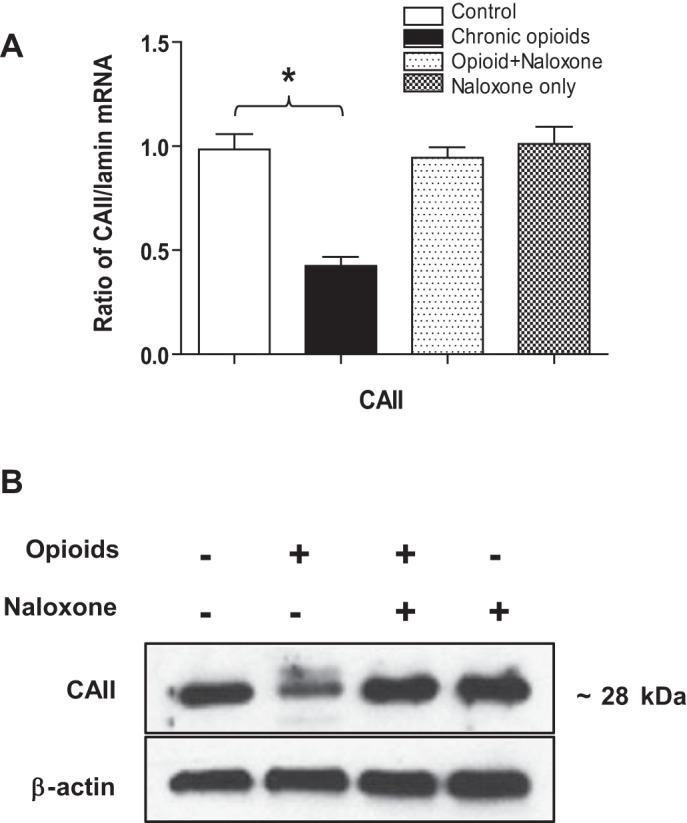
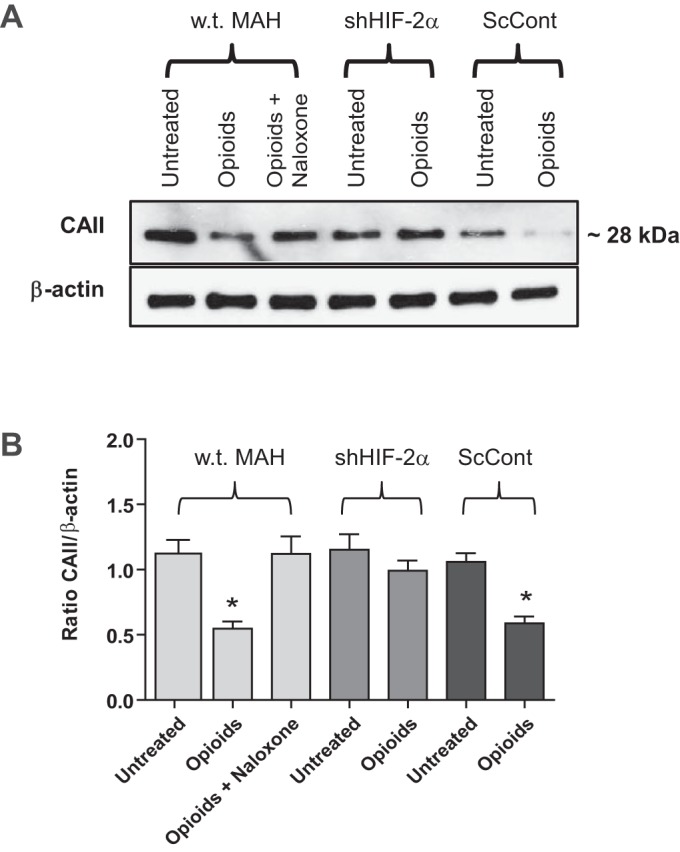
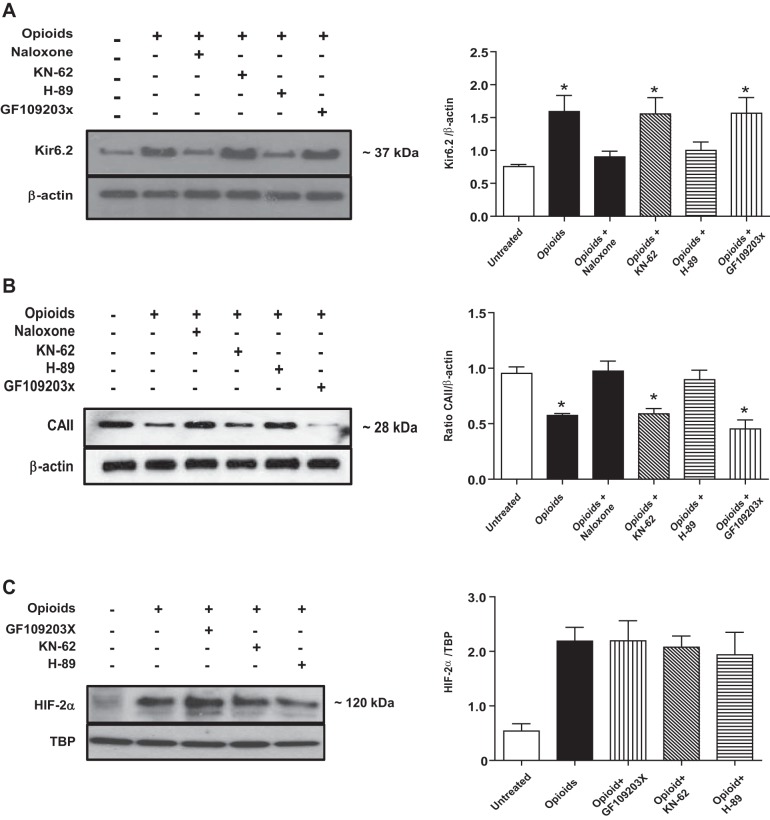
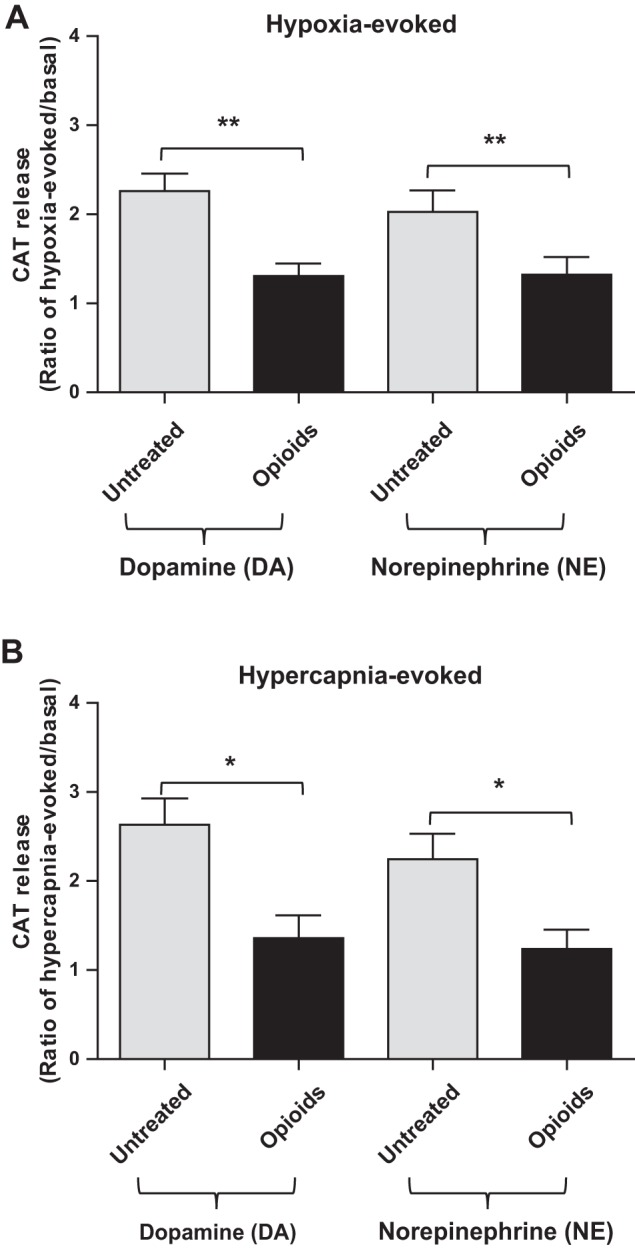
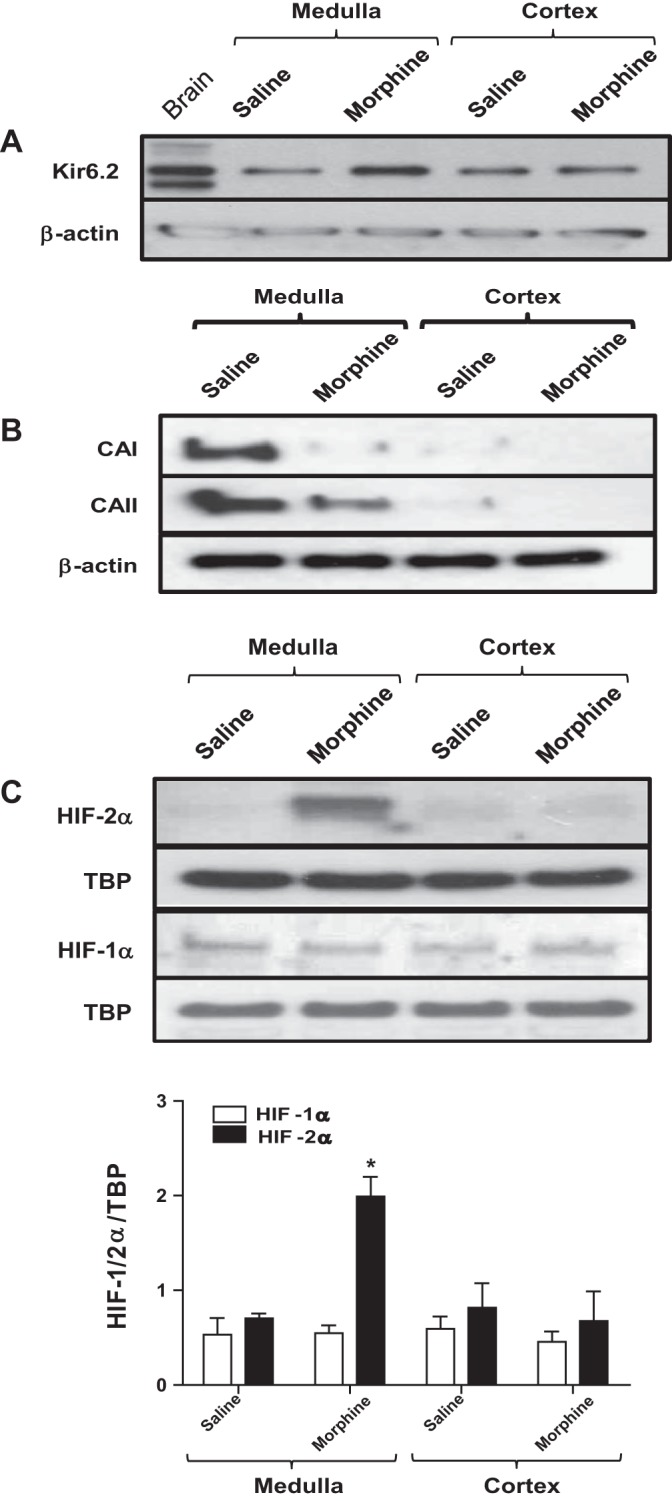
At birth, asphyxial stressors such as hypoxia and hypercapnia are important physiological stimuli for adrenal catecholamine release that is critical for the proper transition to extrauterine life. We recently showed that chronic opioids blunt chemosensitivity of neonatal rat adrenomedullary chromaffin cells (AMCs) to hypoxia and hypercapnia. This blunting was attributable to increased ATP-sensitive K(+) (KATP) channel and decreased carbonic anhydrase (CA) I and II expression, respectively, and involved μ- and δ-opioid receptor signaling pathways. To address underlying molecular mechanisms, we first exposed an O2- and CO2-sensitive, immortalized rat chromaffin cell line (MAH cells) to combined μ {[d-Arg(2),Ly(4)]dermorphin-(1-4)-amide}- and δ ([d-Pen(2),5,P-Cl-Phe(4)]enkephalin)-opioid agonists (2 μM) for ∼7 days. Western blot and quantitative real-time PCR analysis revealed that chronic opioids increased KATP channel subunit Kir6.2 and decreased CAII expression; both effects were blocked by naloxone and were absent in hypoxia-inducible factor (HIF)-2α-deficient MAH cells. Chronic opioids also stimulated HIF-2α accumulation along a time course similar to Kir6.2. Chromatin immunoprecipitation assays on opioid-treated cells revealed the binding of HIF-2α to a hypoxia response element in the promoter region of the Kir6.2 gene. The opioid-induced regulation of Kir6.2 and CAII was dependent on protein kinase A, but not protein kinase C or calmodulin kinase, activity. Interestingly, a similar pattern of HIF-2α, Kir6.2, and CAII regulation (including downregulation of CAI) was replicated in chromaffin tissue obtained from rat pups born to dams exposed to morphine throughout gestation. Collectively, these data reveal novel mechanisms by which chronic opioids blunt asphyxial chemosensitivity in AMCs, thereby contributing to abnormal arousal responses in the offspring of opiate-addicted mothers.
Keywords: adenosine 5′-triphosphate-sensitive potassium channels; carbonic anhydrase; hypoxia-inducible factor-2α; opioid.
Copyright © 2014 the American Physiological Society.
Figures
References
-
- Bishop T, Gallagher D, Pascual A, Lygate CA, de Bono JP, Nicholls LG, Ortega-Saenz P, Oster H, Wijeyekoon B, Sutherland AI, Grosfeld A, Aragones J, Schneider M, van Geyte K, Teixeira D, Diez-Juan A, Lopez-Barneo J, Channon KM, Maxwell PH, Pugh CW, Davies AM, Carmeliet P, Ratcliffe PJ. Abnormal sympathoadrenal development and systemic hypotension in PHD3−/− mice. Mol Cell Biol 28: 3386–3400, 2008 - PMC - PubMed
-
- Brown ST, Kelly KF, Daniel JM, Nurse CA. Hypoxia inducible factor (HIF)-2 alpha is required for the development of the catecholaminergic phenotype of sympathoadrenal cells. J Neurochem 110: 622–630, 2009 - PubMed
-
- Brown ST, Nurse CA. Induction of HIF-2alpha is dependent on mitochondrial O2 consumption in an O2-sensitive adrenomedullary chromaffin cell line. Am J Physiol Cell Physiol 294: C1305–C1312, 2008 - PubMed
-
- Burns L, Conroy E, Mattick RP. Infant mortality among women on a methadone program during pregnancy. Drug Alcohol Rev 29: 551–556, 2010 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials