

Phytochemicals perturb membranes and promiscuously alter protein function
- PMID: 24901212
- PMCID: PMC4136704
- DOI: 10.1021/cb500086e
Phytochemicals perturb membranes and promiscuously alter protein function
Abstract
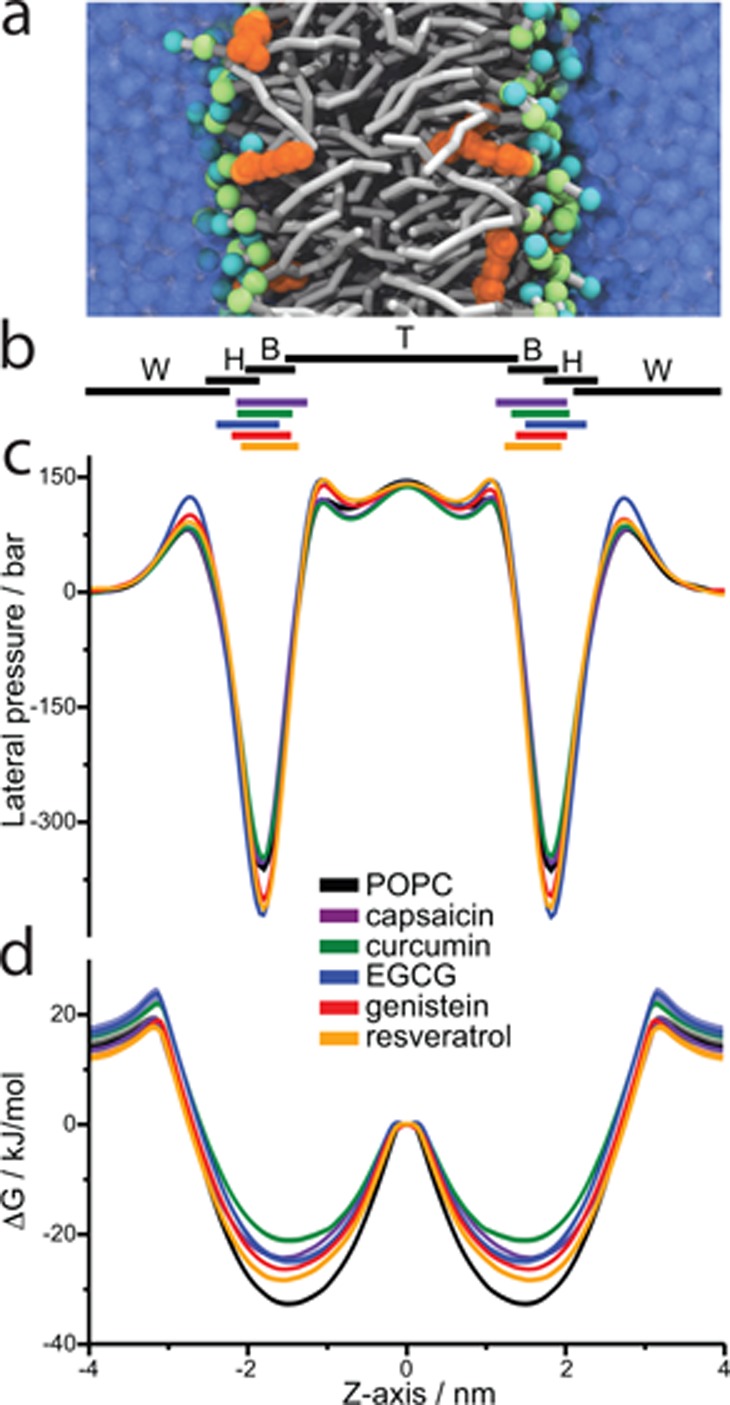
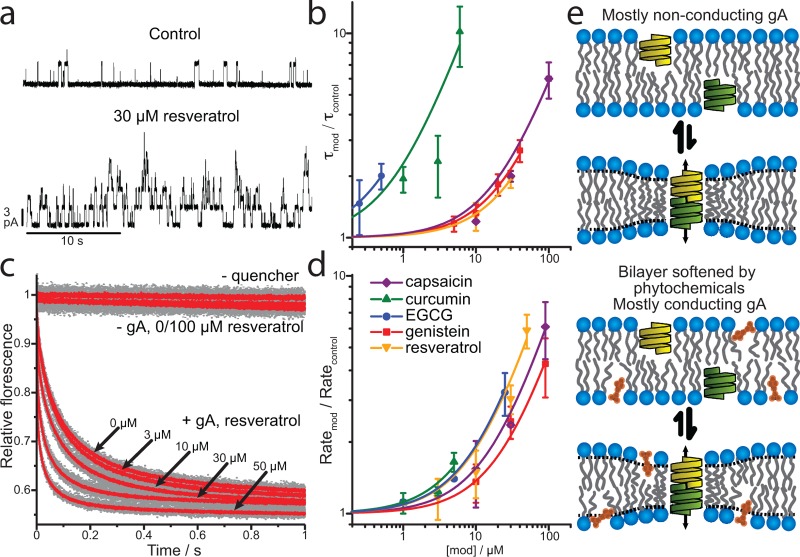
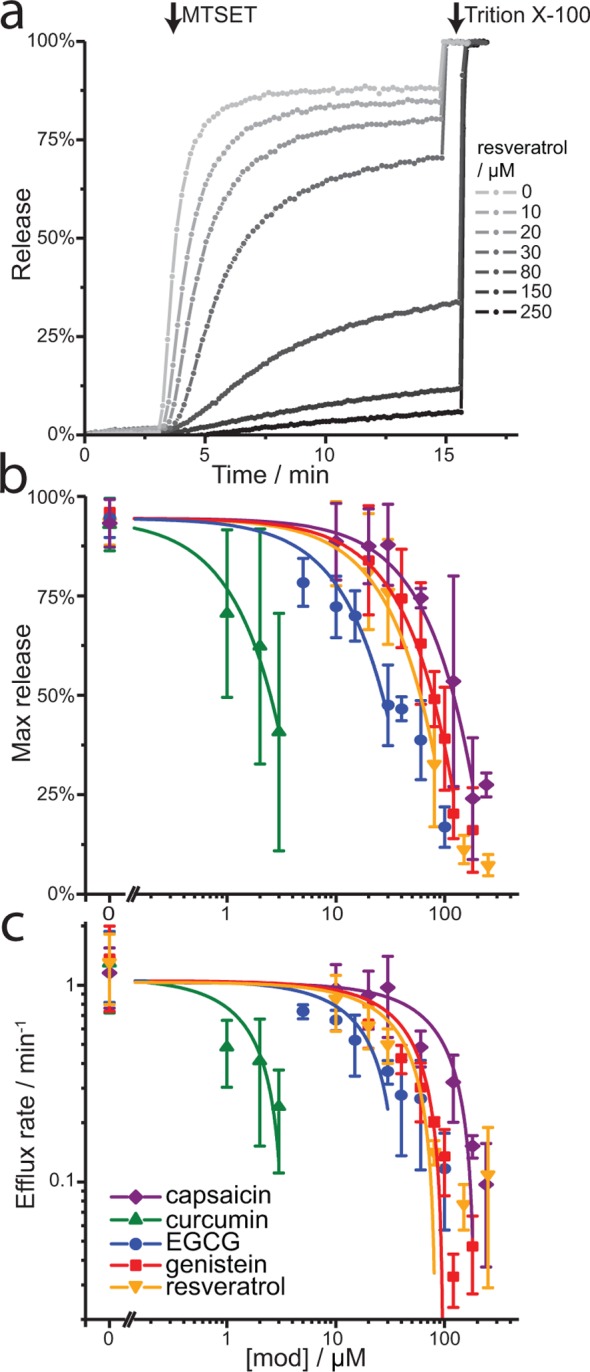
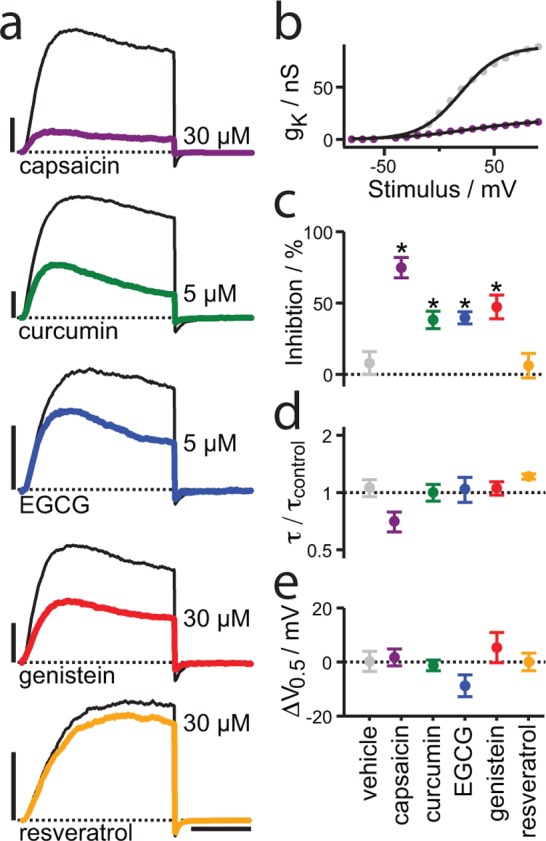
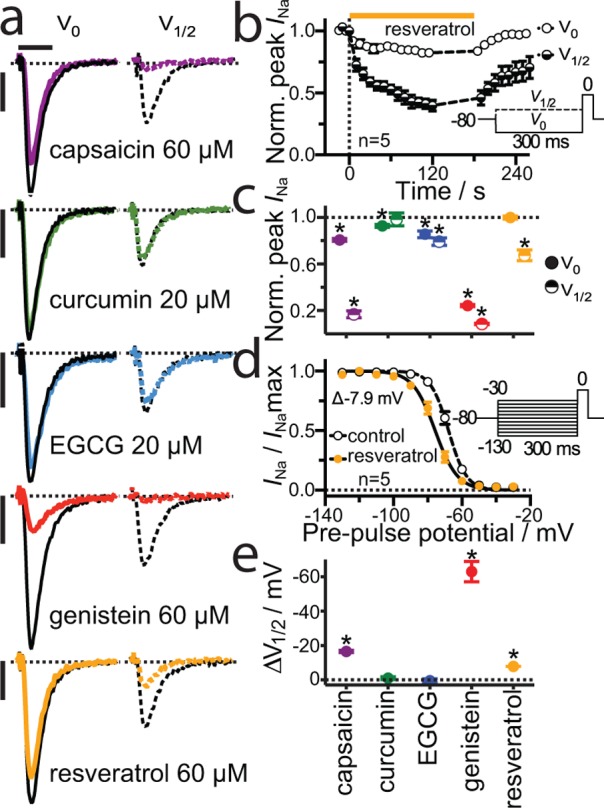
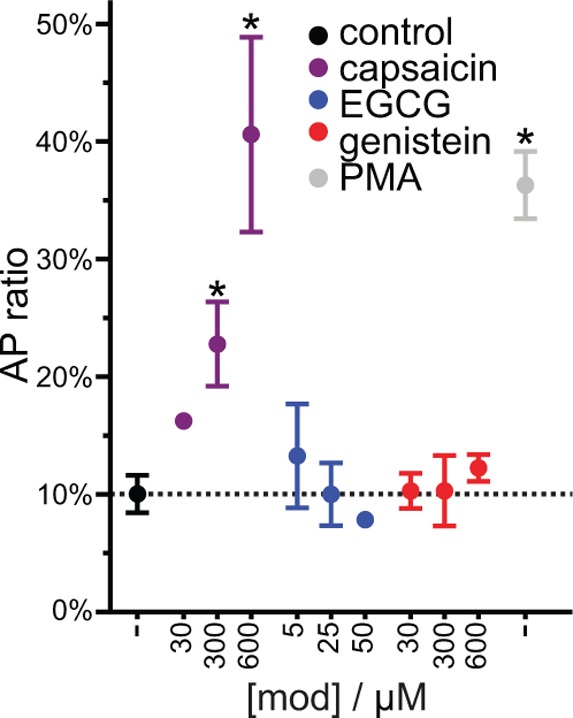
A wide variety of phytochemicals are consumed for their perceived health benefits. Many of these phytochemicals have been found to alter numerous cell functions, but the mechanisms underlying their biological activity tend to be poorly understood. Phenolic phytochemicals are particularly promiscuous modifiers of membrane protein function, suggesting that some of their actions may be due to a common, membrane bilayer-mediated mechanism. To test whether bilayer perturbation may underlie this diversity of actions, we examined five bioactive phenols reported to have medicinal value: capsaicin from chili peppers, curcumin from turmeric, EGCG from green tea, genistein from soybeans, and resveratrol from grapes. We find that each of these widely consumed phytochemicals alters lipid bilayer properties and the function of diverse membrane proteins. Molecular dynamics simulations show that these phytochemicals modify bilayer properties by localizing to the bilayer/solution interface. Bilayer-modifying propensity was verified using a gramicidin-based assay, and indiscriminate modulation of membrane protein function was demonstrated using four proteins: membrane-anchored metalloproteases, mechanosensitive ion channels, and voltage-dependent potassium and sodium channels. Each protein exhibited similar responses to multiple phytochemicals, consistent with a common, bilayer-mediated mechanism. Our results suggest that many effects of amphiphilic phytochemicals are due to cell membrane perturbations, rather than specific protein binding.
Figures
References
-
- Joe B.; Vijaykumar M.; Lokesh B. R. (2004) Biological properties of curcumin—cellular and molecular mechanisms of action. Crit. Rev. Food Sci. Nutr. 44, 97–111. - PubMed
-
- Shishodia S.; Sethi G.; Aggarwal B. B. (2005) Curcumin: Getting back to the roots. Ann. N.Y. Acad. Sci. 1056, 206–217. - PubMed
-
- Shimizu M.; Weinstein I. B. (2005) Modulation of signal transduction by tea catechins and related phytochemicals. Mutat. Res. 591, 147–160. - PubMed
-
- Khan N.; Afaq F.; Saleem M.; Ahmad N.; Mukhtar H. (2006) Targeting multiple signaling pathways by green tea polyphenol (−)-epigallocatechin-3-gallate. Cancer Res. 66, 2500–2505. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
