

A normative theory of forgetting: lessons from the fruit fly
- PMID: 24901935
- PMCID: PMC4046926
- DOI: 10.1371/journal.pcbi.1003640
A normative theory of forgetting: lessons from the fruit fly
Abstract
Recent experiments revealed that the fruit fly Drosophila melanogaster has a dedicated mechanism for forgetting: blocking the G-protein Rac leads to slower and activating Rac to faster forgetting. This active form of forgetting lacks a satisfactory functional explanation. We investigated optimal decision making for an agent adapting to a stochastic environment where a stimulus may switch between being indicative of reward or punishment. Like Drosophila, an optimal agent shows forgetting with a rate that is linked to the time scale of changes in the environment. Moreover, to reduce the odds of missing future reward, an optimal agent may trade the risk of immediate pain for information gain and thus forget faster after aversive conditioning. A simple neuronal network reproduces these features. Our theory shows that forgetting in Drosophila appears as an optimal adaptive behavior in a changing environment. This is in line with the view that forgetting is adaptive rather than a consequence of limitations of the memory system.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
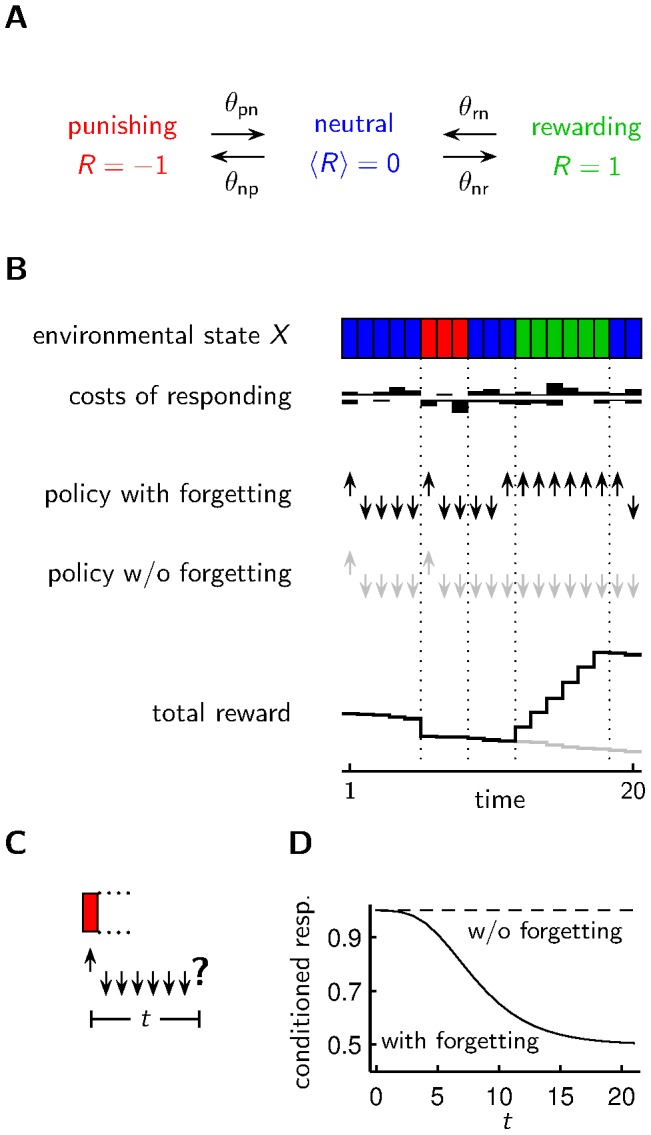
 between being rewarding, neutral or punishing. Unless mentioned otherwise, we choose
between being rewarding, neutral or punishing. Unless mentioned otherwise, we choose  and
and  . B Based on a policy (with forgetting, without forgetting) which may depend on past observations of the environmental state and current costs of responding, an agent shows the appetitive reaction (upward arrow) or the aversive reaction (downward arrow). The stochastic costs (i.i.d. with an exponential distribution with scale parameter
. B Based on a policy (with forgetting, without forgetting) which may depend on past observations of the environmental state and current costs of responding, an agent shows the appetitive reaction (upward arrow) or the aversive reaction (downward arrow). The stochastic costs (i.i.d. with an exponential distribution with scale parameter  ) for the appetitive/aversive reaction are shown above/below the white line. An agent with a policy that involves forgetting accumulates more reward than an agent without forgetting or immediate forgetting. C In an emulation of a classical conditioning experiment, the agent experiences a defined environmental state, and after a waiting period of length
) for the appetitive/aversive reaction are shown above/below the white line. An agent with a policy that involves forgetting accumulates more reward than an agent without forgetting or immediate forgetting. C In an emulation of a classical conditioning experiment, the agent experiences a defined environmental state, and after a waiting period of length  the agent has to react according to the internal policy. D Different policies lead to different outcomes in classical conditioning experiments. Shown is the fraction of agents choosing the conditioned response (conditioned resp.) at time
the agent has to react according to the internal policy. D Different policies lead to different outcomes in classical conditioning experiments. Shown is the fraction of agents choosing the conditioned response (conditioned resp.) at time  after conditioning for agents subject to individual costs of responding.
after conditioning for agents subject to individual costs of responding.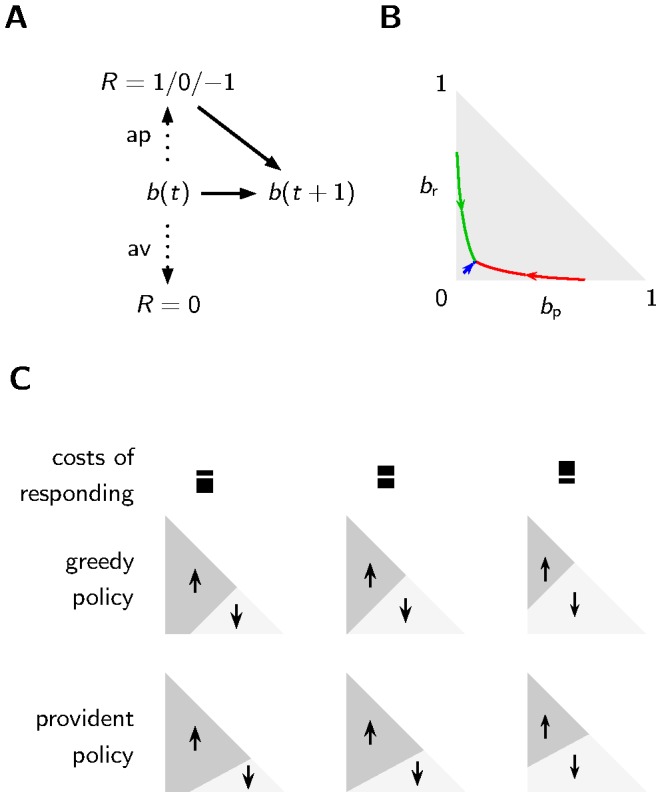
 may influence the choice of the appetitive or aversive reaction. Only after the appetitive reaction, the agent gains new information about the true state of the environment. The belief
may influence the choice of the appetitive or aversive reaction. Only after the appetitive reaction, the agent gains new information about the true state of the environment. The belief  and the agents knowledge about the transition probabilities of the environmental state combined with potentially new information determines the new belief
and the agents knowledge about the transition probabilities of the environmental state combined with potentially new information determines the new belief  . B The starting point of the arrows is a belief found by choosing the appetitive reaction once and receiving reward (green), punishment (red) or no reinforcement (blue). If the agent always chooses the aversive reaction thereafter, the belief drifts to the stationary state along the trajectories shown by the arrows. Possible belief states
. B The starting point of the arrows is a belief found by choosing the appetitive reaction once and receiving reward (green), punishment (red) or no reinforcement (blue). If the agent always chooses the aversive reaction thereafter, the belief drifts to the stationary state along the trajectories shown by the arrows. Possible belief states  with
with  can be represented as a point in the “belief space” (gray shaded triangle). C The regions in the belief space favoring the appetitive reaction (dark shading, upward arrow) over the aversive reaction (bright shading, downward arrow) depend on the policy and the costs of responding. The provident policy (lowest row) is biased towards the appetitive reaction. A larger cost for the aversive reaction than for the appetitive reaction (left column) decreases the region of the aversive reaction.
can be represented as a point in the “belief space” (gray shaded triangle). C The regions in the belief space favoring the appetitive reaction (dark shading, upward arrow) over the aversive reaction (bright shading, downward arrow) depend on the policy and the costs of responding. The provident policy (lowest row) is biased towards the appetitive reaction. A larger cost for the aversive reaction than for the appetitive reaction (left column) decreases the region of the aversive reaction.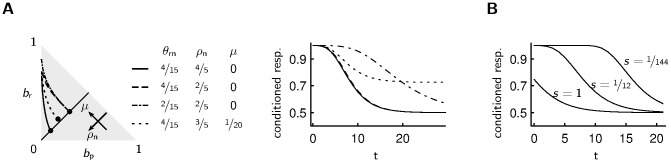
 or increasing average reward
or increasing average reward  . How fast the belief drifts towards the stationary state after receiving reward depends on the parameter
. How fast the belief drifts towards the stationary state after receiving reward depends on the parameter  that controls the “timescale of changes”. Right: Changing the probability of the neutral state
that controls the “timescale of changes”. Right: Changing the probability of the neutral state  only marginally affects the forgetting curve (solid and dashed line). A smaller rate of changes
only marginally affects the forgetting curve (solid and dashed line). A smaller rate of changes  leads to slower forgetting (dash-dotted curve). A positive average reward
leads to slower forgetting (dash-dotted curve). A positive average reward  leads to a higher fraction of agents choosing the appetitive reaction, which is here the conditioned response (dotted curve). B For a large variance of costs of responding (curve with scale parameter of the exponential distribution
leads to a higher fraction of agents choosing the appetitive reaction, which is here the conditioned response (dotted curve). B For a large variance of costs of responding (curve with scale parameter of the exponential distribution  ) there are some agents that do not exhibit the conditioned response immediately after conditioning, since the costs of the conditioned response are too large. If the variance of the costs of responding is small (curve with
) there are some agents that do not exhibit the conditioned response immediately after conditioning, since the costs of the conditioned response are too large. If the variance of the costs of responding is small (curve with  ), most agents choose the conditioned response until their belief gets close to the stationary belief state.
), most agents choose the conditioned response until their belief gets close to the stationary belief state.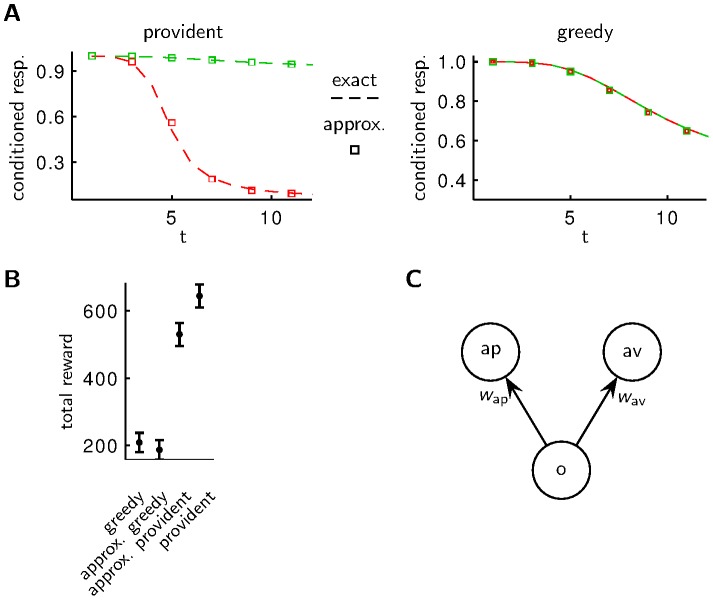
 time bins (compare to Fig. 1B) is larger for the provident policy than for the greedy policy. Plotted are mean and s.e.m. for 40 trials. C Similar performances are obtained with a simple, approximative implementation of the optimal strategy with synaptic strengths
time bins (compare to Fig. 1B) is larger for the provident policy than for the greedy policy. Plotted are mean and s.e.m. for 40 trials. C Similar performances are obtained with a simple, approximative implementation of the optimal strategy with synaptic strengths  and
and  connecting an odor detecting neuron (o) to action neurons “approach” (ap) and “avoid” (av). In the absence of any stimulus (odor) the synaptic strengths decay with different time constants for the approximative provident policy and with the same time constants for the approximative greedy policy. When an odor is present, the synaptic strengths change in a Hebbian way in the case of reward and in an anti-Hebbian way in the case of punishment, i.e.
connecting an odor detecting neuron (o) to action neurons “approach” (ap) and “avoid” (av). In the absence of any stimulus (odor) the synaptic strengths decay with different time constants for the approximative provident policy and with the same time constants for the approximative greedy policy. When an odor is present, the synaptic strengths change in a Hebbian way in the case of reward and in an anti-Hebbian way in the case of punishment, i.e.  /
/ increase/decrease for reward and decrease/increase for punishment.
increase/decrease for reward and decrease/increase for punishment.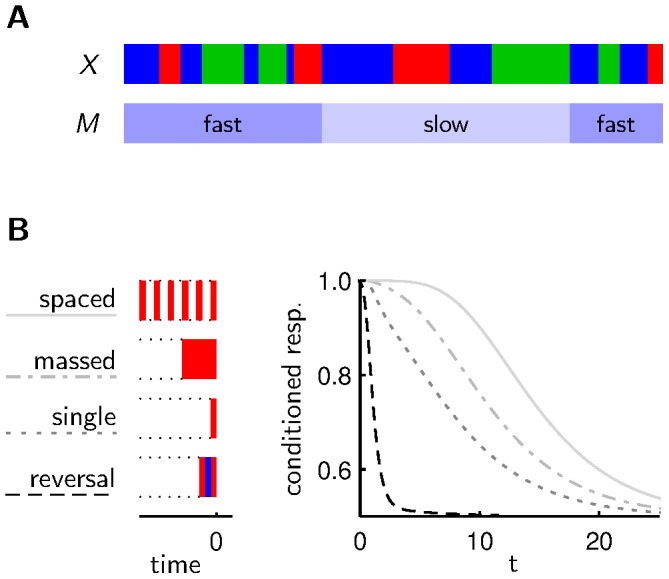
 , which can be in a slow or fast state. B As observed in experiments with Drosophila, our model agents show slowest forgetting after spaced training and fastest forgetting of the last association after reversal training. In our model, this result appears as a consequence of spaced training being most informative about slow transitions, whereas reversal training is most informative about fast transitions.
, which can be in a slow or fast state. B As observed in experiments with Drosophila, our model agents show slowest forgetting after spaced training and fastest forgetting of the last association after reversal training. In our model, this result appears as a consequence of spaced training being most informative about slow transitions, whereas reversal training is most informative about fast transitions.References
-
- Tully T, Quinn W (1985) Classical conditioning and retention in normal and mutant Drosophila melanogaster. Journal of Comparative Physiology A 157: 263–277. - PubMed
-
- Rosenzweig ES, Barnes Ca, McNaughton BL (2002) Making room for new memories. Nature neuroscience 5: 6–8. - PubMed
-
- Storm BC (2011) The benefit of forgetting in thinking and remembering. Current Directions in Psychological Science 20: 291–295.
-
- Wixted JT (2004) The Psychology and Neuroscience of Forgetting. Annual review of psychology 55: 235–69. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
