

Alternative polyadenylation in the nervous system: to what lengths will 3' UTR extensions take us?
- PMID: 24903459
- PMCID: PMC4503322
- DOI: 10.1002/bies.201300174
Alternative polyadenylation in the nervous system: to what lengths will 3' UTR extensions take us?
Abstract
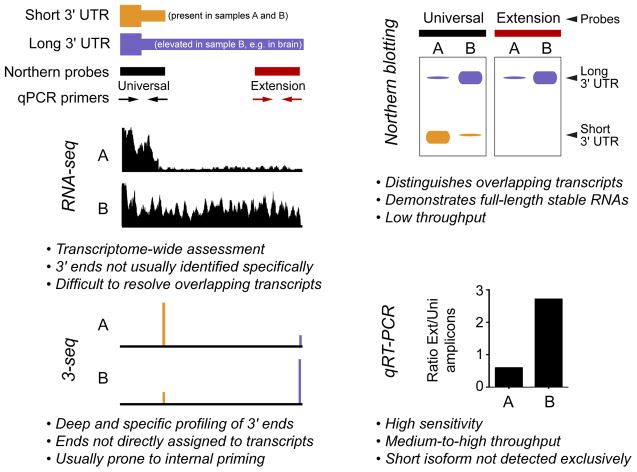
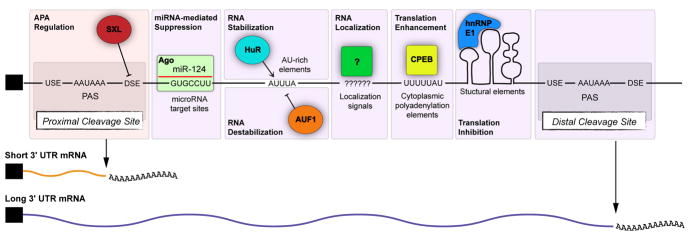
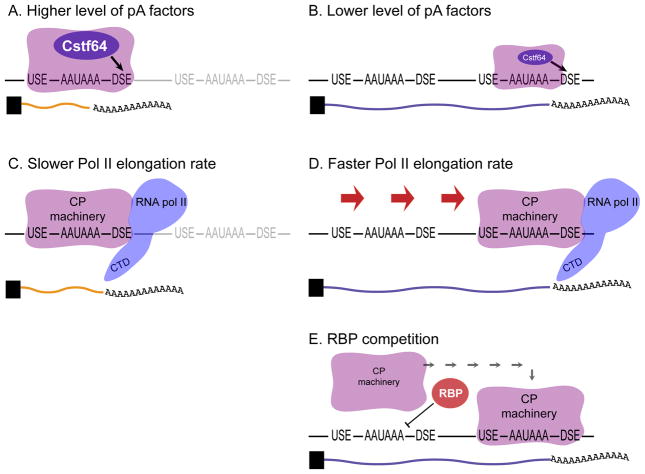
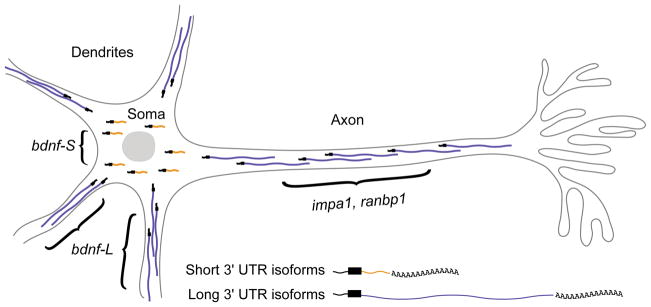
Alternative cleavage and polyadenylation (APA) can diversify coding and non-coding regions, but has particular impact on increasing 3' UTR diversity. Through the gain or loss of regulatory elements such as RNA binding protein and microRNA sites, APA can influence transcript stability, localization, and translational efficiency. Strikingly, the central nervous systems of invertebrate and vertebrate species express a broad range of transcript isoforms bearing extended 3' UTRs. The molecular mechanism that permits proximal 3' end bypass in neurons is mysterious, and only beginning to be elucidated. This landscape of neural 3' UTR extensions, many reaching unprecedented lengths, may help service the unique post-transcriptional regulatory needs of neurons. A combination of approaches, including transcriptome-wide profiling, genetic screening to identify APA factors, biochemical dissection of alternative 3' end formation, and manipulation of individual neural APA targets, will be necessary to gain fuller perspectives on the mechanism and biology of neural-specific 3' UTR lengthening.
Keywords: 3′ UTR; RNA-binding protein; alternative polyadenylation; miRNA; nervous system.
© 2014 WILEY Periodicals, Inc.
Figures
References
-
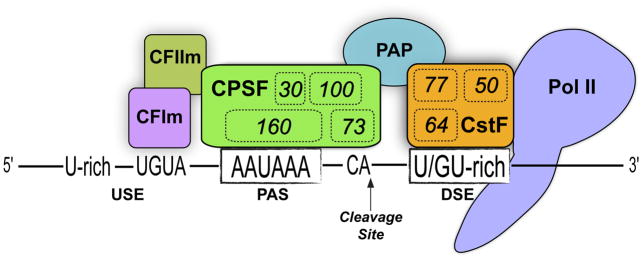
- Colgan DF, Manley JL. Mechanism and regulation of mRNA polyadenylation. Genes & development. 1997;11:2755–66. - PubMed
-
- Martin G, Gruber AR, Keller W, Zavolan M. Genome-wide analysis of pre-mRNA 3′ end processing reveals a decisive role of human cleavage factor I in the regulation of 3′ UTR length. Cell reports. 2012;1:753–63. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
