

Recombination signal binding protein for Ig-κJ region regulates juxtaglomerular cell phenotype by activating the myo-endocrine program and suppressing ectopic gene expression
- PMID: 24904090
- PMCID: PMC4279731
- DOI: 10.1681/ASN.2013101045
Recombination signal binding protein for Ig-κJ region regulates juxtaglomerular cell phenotype by activating the myo-endocrine program and suppressing ectopic gene expression
Abstract
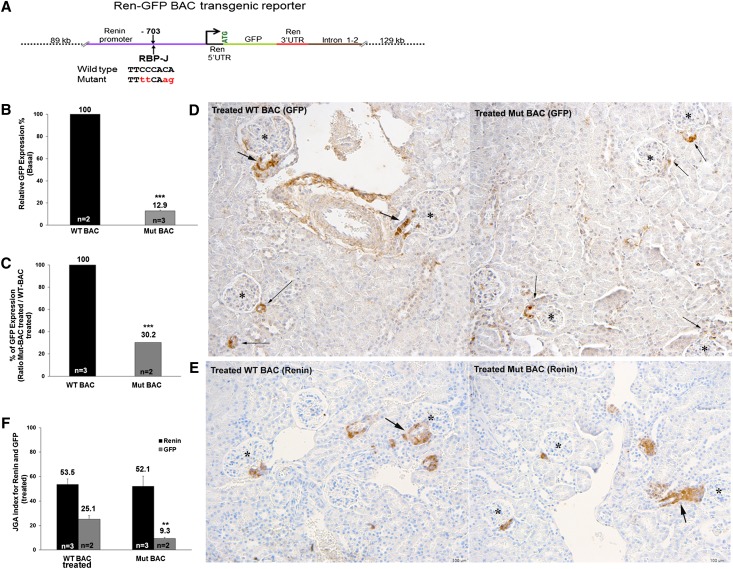
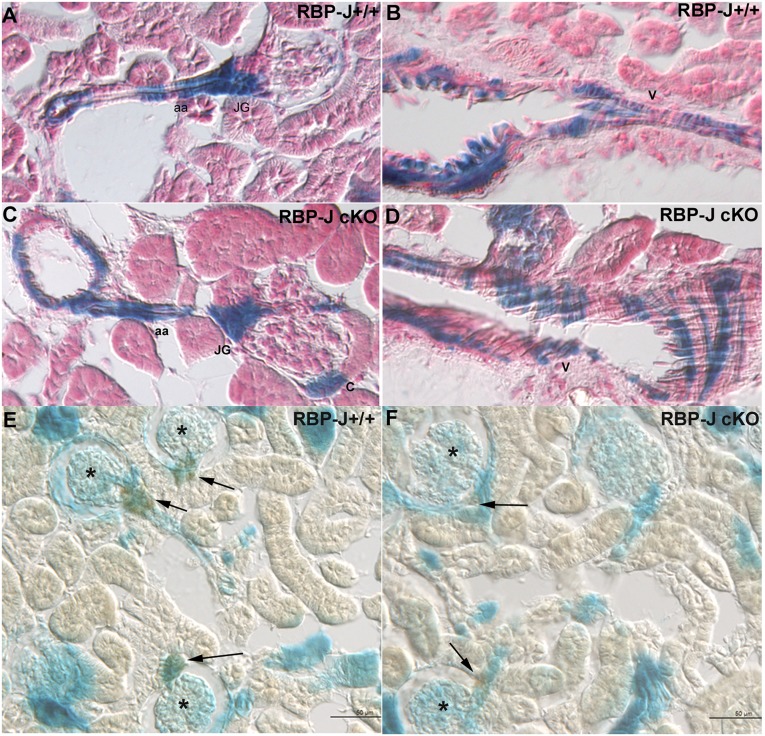
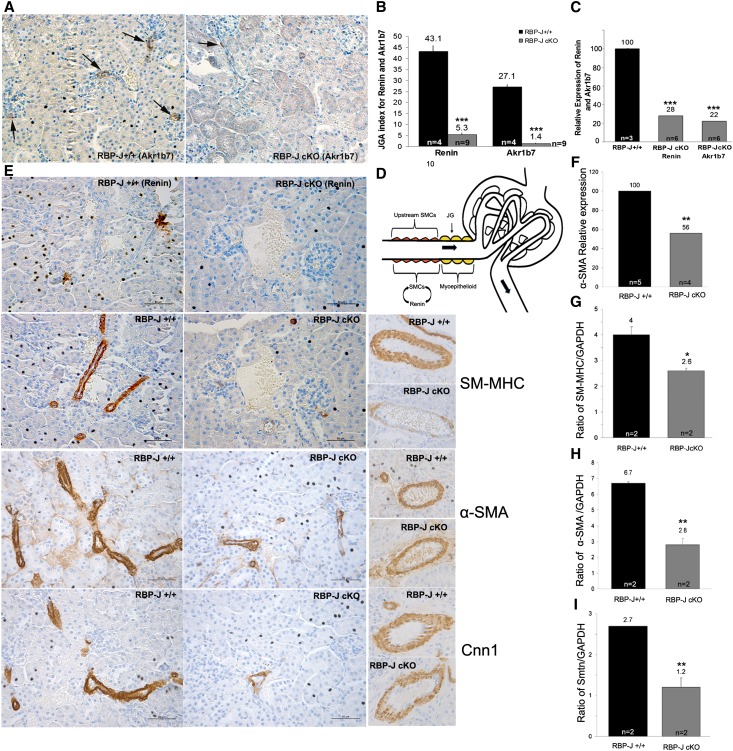
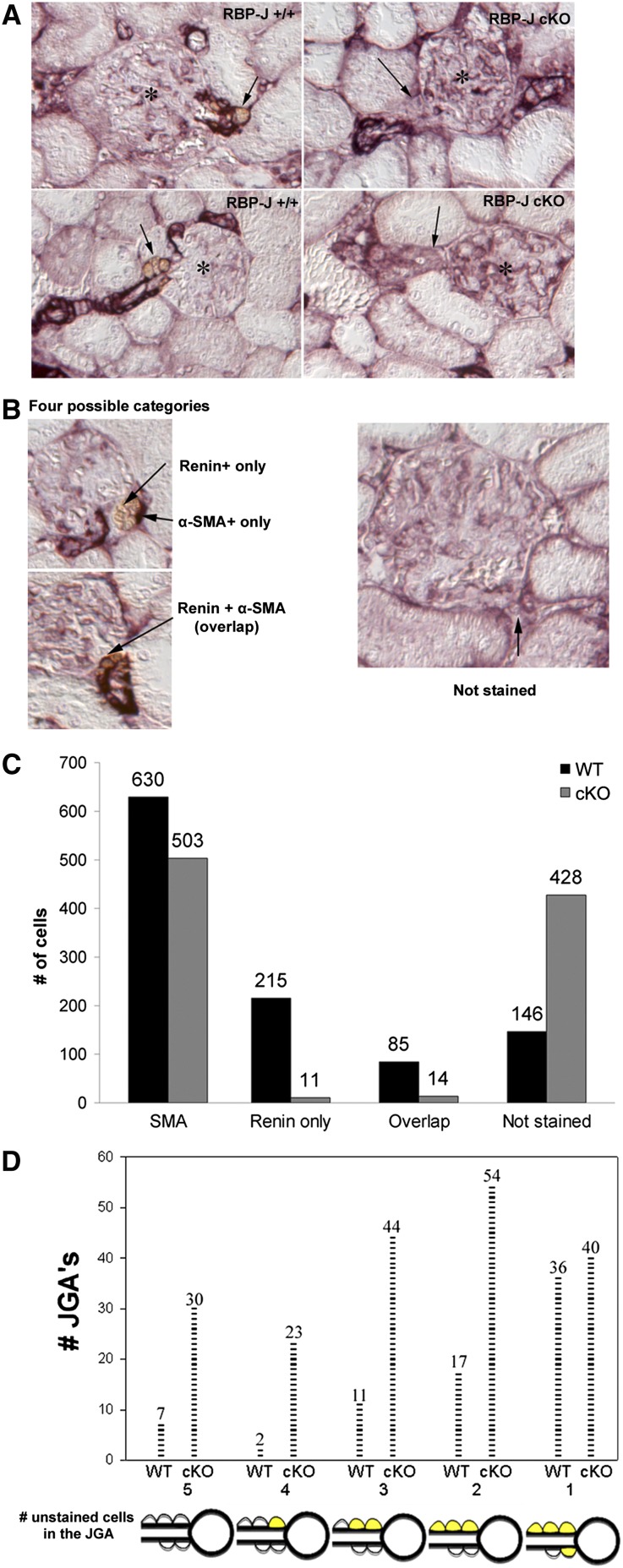
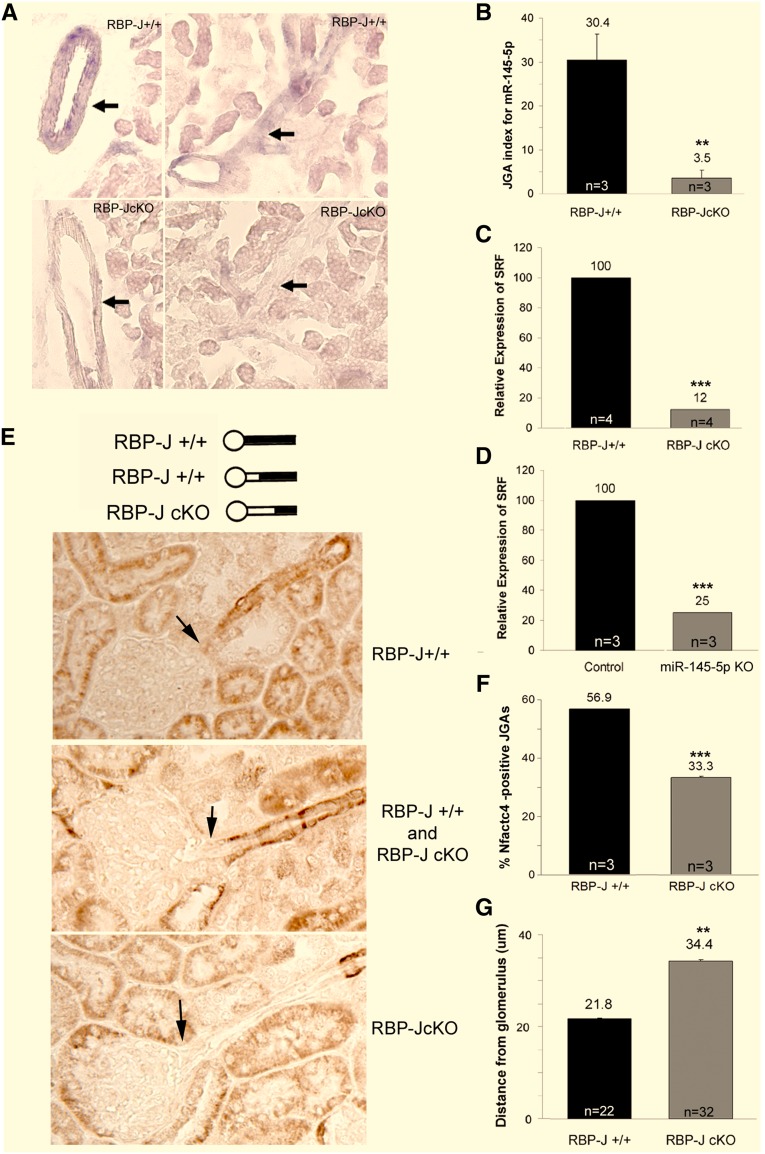
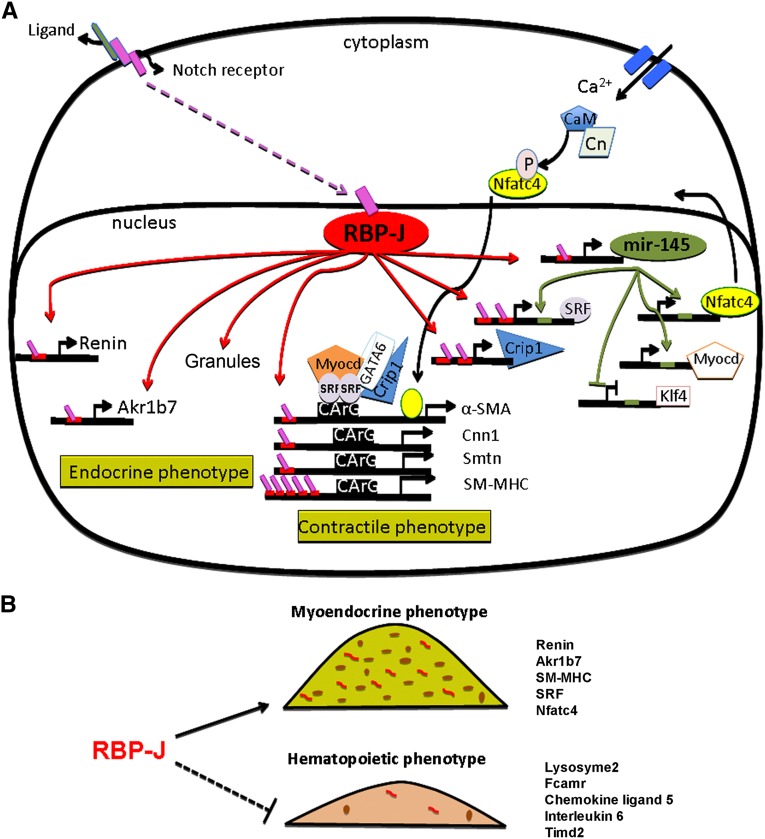
Recombination signal binding protein for Ig-κJ region (RBP-J), the major downstream effector of Notch signaling, is necessary to maintain the number of renin-positive juxtaglomerular cells and the plasticity of arteriolar smooth muscle cells to re-express renin when homeostasis is threatened. We hypothesized that RBP-J controls a repertoire of genes that defines the phenotype of the renin cell. Mice bearing a bacterial artificial chromosome reporter with a mutated RBP-J binding site in the renin promoter had markedly reduced reporter expression at the basal state and in response to a homeostatic challenge. Mice with conditional deletion of RBP-J in renin cells had decreased expression of endocrine (renin and Akr1b7) and smooth muscle (Acta2, Myh11, Cnn1, and Smtn) genes and regulators of smooth muscle expression (miR-145, SRF, Nfatc4, and Crip1). To determine whether RBP-J deletion decreased the endowment of renin cells, we traced the fate of these cells in RBP-J conditional deletion mice. Notably, the lineage staining patterns in mutant and control kidneys were identical, although mutant kidneys had fewer or no renin-expressing cells in the juxtaglomerular apparatus. Microarray analysis of mutant arterioles revealed upregulation of genes usually expressed in hematopoietic cells. Thus, these results suggest that RBP-J maintains the identity of the renin cell by not only activating genes characteristic of the myo-endocrine phenotype but also, preventing ectopic gene expression and adoption of an aberrant phenotype, which could have severe consequences for the control of homeostasis.
Keywords: cell fate; hematopoiesis; juxtaglomerular cell; renin; smooth muscle.
Copyright © 2015 by the American Society of Nephrology.
Figures

References
-
- Gomez RA, Chevalier RL, Sturgill BC, Johns DW, Peach MJ, Carey RM: Maturation of the intrarenal renin distribution in Wistar-Kyoto rats. J Hypertens 4: S31–S33, 1986
-
- Gomez RA, Lynch KR, Chevalier RL, Wilfong N, Everett A, Carey RM, Peach MJ: Renin and angiotensinogen gene expression in maturing rat kidney. Am J Physiol 254: F582–F587, 1988 - PubMed
-
- Gomez RA, Chevalier RL, Everett AD, Elwood JP, Peach MJ, Lynch KR, Carey RM: Recruitment of renin gene-expressing cells in adult rat kidneys. Am J Physiol 259: F660–F665, 1990 - PubMed
-
- Sequeira Lopez ML, Pentz ES, Robert B, Abrahamson DR, Gomez RA: Embryonic origin and lineage of juxtaglomerular cells. Am J Physiol Renal Physiol 281: F345–F356, 2001 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
