

Modelling mammalian cellular quiescence
- PMID: 24904737
- PMCID: PMC3996586
- DOI: 10.1098/rsfs.2013.0074
Modelling mammalian cellular quiescence
Abstract
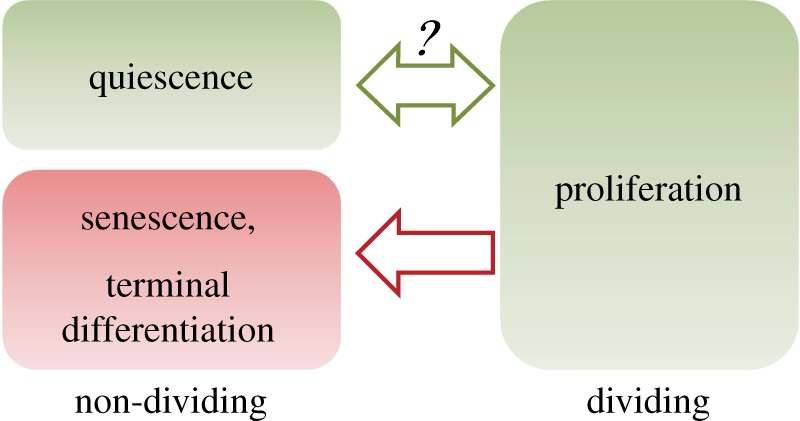
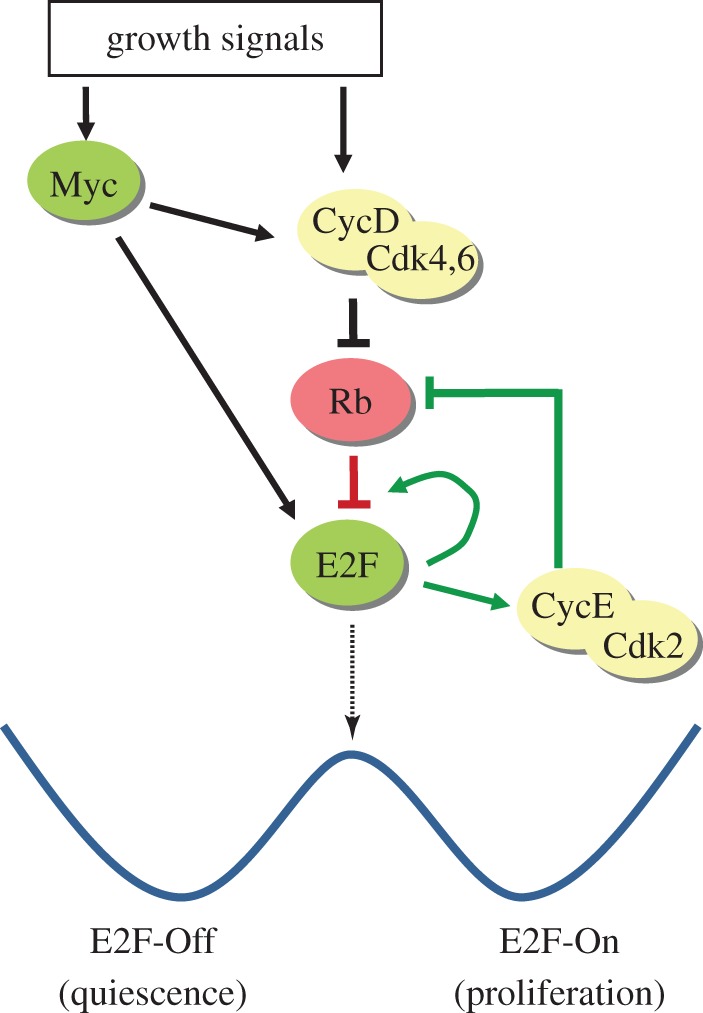
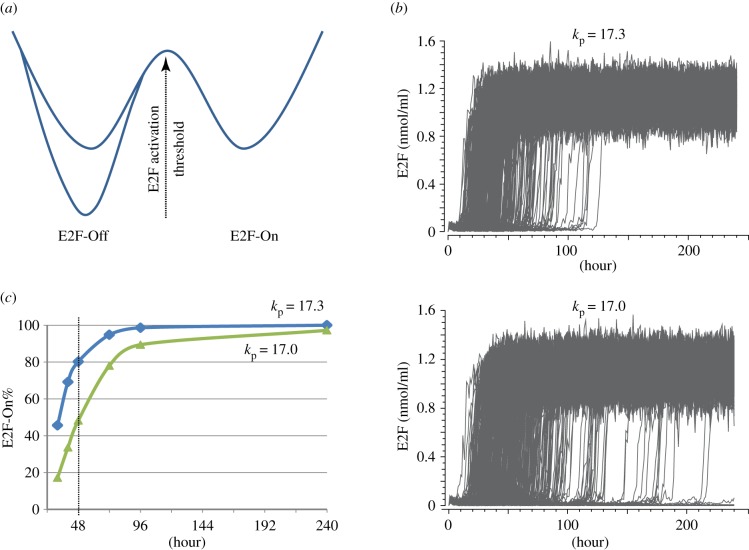
Cellular quiescence is a reversible non-proliferating state. The reactivation of 'sleep-like' quiescent cells (e.g. fibroblasts, lymphocytes and stem cells) into proliferation is crucial for tissue repair and regeneration and a key to the growth, development and health of higher multicellular organisms, such as mammals. Quiescence has been a primarily phenotypic description (i.e. non-permanent cell cycle arrest) and poorly studied. However, contrary to the earlier thinking that quiescence is simply a passive and dormant state lacking proliferating activities, recent studies have revealed that cellular quiescence is actively maintained in the cell and that it corresponds to a collection of heterogeneous states. Recent modelling and experimental work have suggested that an Rb-E2F bistable switch plays a pivotal role in controlling the quiescence-proliferation balance and the heterogeneous quiescent states. Other quiescence regulatory activities may crosstalk with and impinge upon the Rb-E2F bistable switch, forming a gene network that controls the cells' quiescent states and their dynamic transitions to proliferation in response to noisy environmental signals. Elucidating the dynamic control mechanisms underlying quiescence may lead to novel therapeutic strategies that re-establish normal quiescent states, in a variety of hyper- and hypo-proliferative diseases, including cancer and ageing.
Keywords: bistable switch; cell cycle; cellular quiescence; gene network.
Figures
References
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources