

Reorganization of enhancer patterns in transition from naive to primed pluripotency
- PMID: 24905168
- PMCID: PMC4491504
- DOI: 10.1016/j.stem.2014.04.003
Reorganization of enhancer patterns in transition from naive to primed pluripotency
Abstract
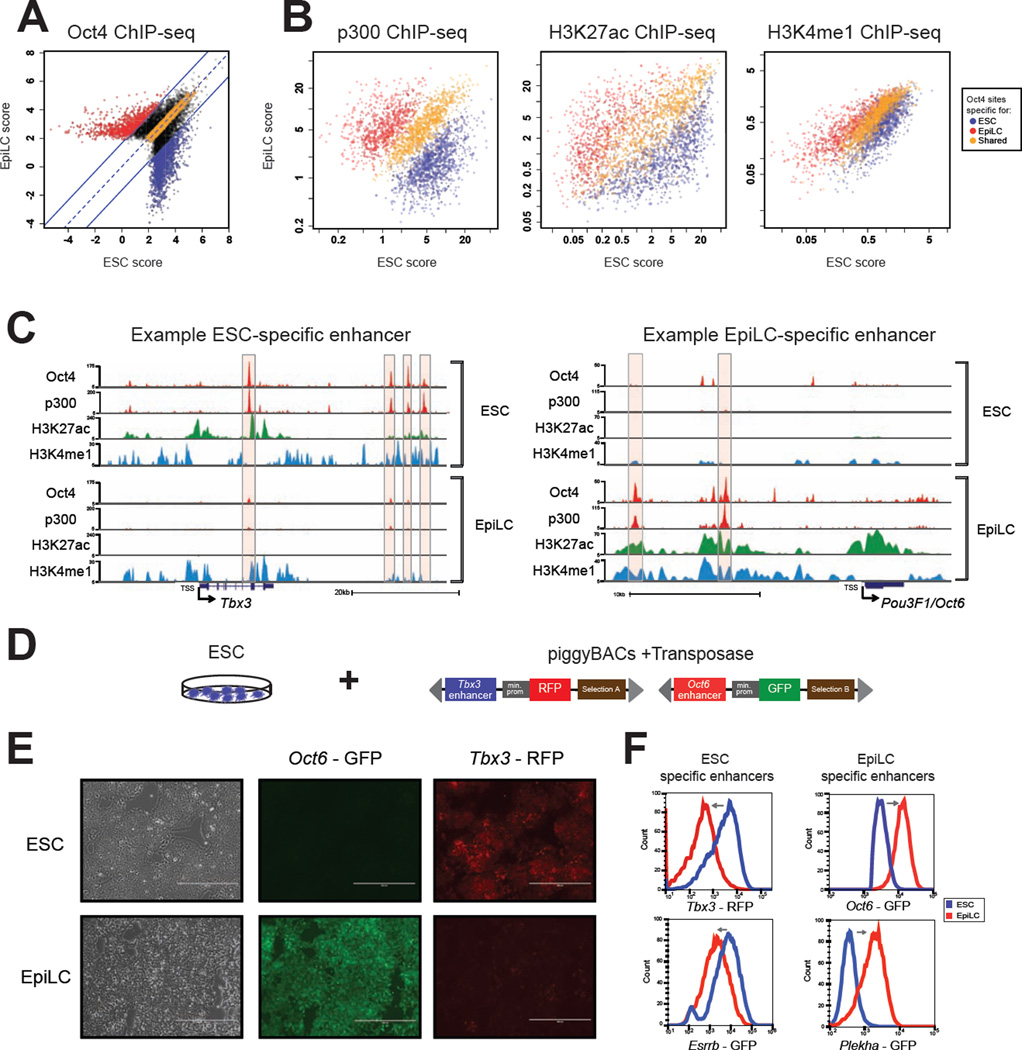
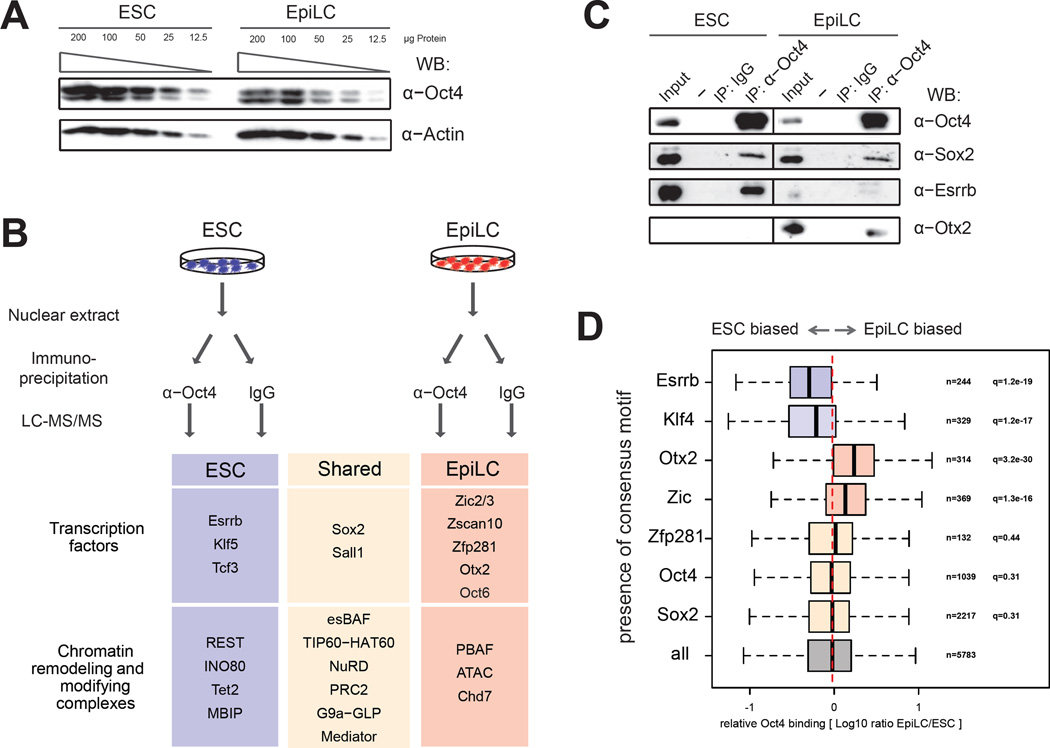
Naive and primed pluripotency is characterized by distinct signaling requirements, transcriptomes, and developmental properties, but both cellular states share key transcriptional regulators: Oct4, Sox2, and Nanog. Here, we demonstrate that transition between these two pluripotent states is associated with widespread Oct4 relocalization, mirrored by global rearrangement of enhancer chromatin landscapes. Our genomic and biochemical analyses identified candidate mediators of primed state-specific Oct4 binding, including Otx2 and Zic2/3. Even when differentiation cues are blocked, premature Otx2 overexpression is sufficient to exit the naive state, induce transcription of a substantial subset of primed pluripotency-associated genes, and redirect Oct4 to previously inaccessible enhancer sites. However, the ability of Otx2 to engage new enhancer regions is determined by its levels, cis-encoded properties of the sites, and the signaling environment. Our results illuminate regulatory mechanisms underlying pluripotency and suggest that the capacity of transcription factors such as Otx2 and Oct4 to pioneer new enhancer sites is highly context dependent.
Copyright © 2014 Elsevier Inc. All rights reserved.
Figures





References
-
- Acampora D, Di Giovannantonio LG, Simeone A. Otx2 is an intrinsic determinant of the embryonic stem cell state and is required for transition to a stable epiblast stem cell condition. Dev. Camb. Engl. 2013;140:43–55. - PubMed
-
- Boiani M, Schöler HR. Regulatory networks in embryo-derived pluripotent stem cells. Nat. Rev. Mol. Cell Biol. 2005;6:872–884. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
