

Epigenomic comparison reveals activation of "seed" enhancers during transition from naive to primed pluripotency
- PMID: 24905169
- PMCID: PMC4149284
- DOI: 10.1016/j.stem.2014.05.005
Epigenomic comparison reveals activation of "seed" enhancers during transition from naive to primed pluripotency
Abstract
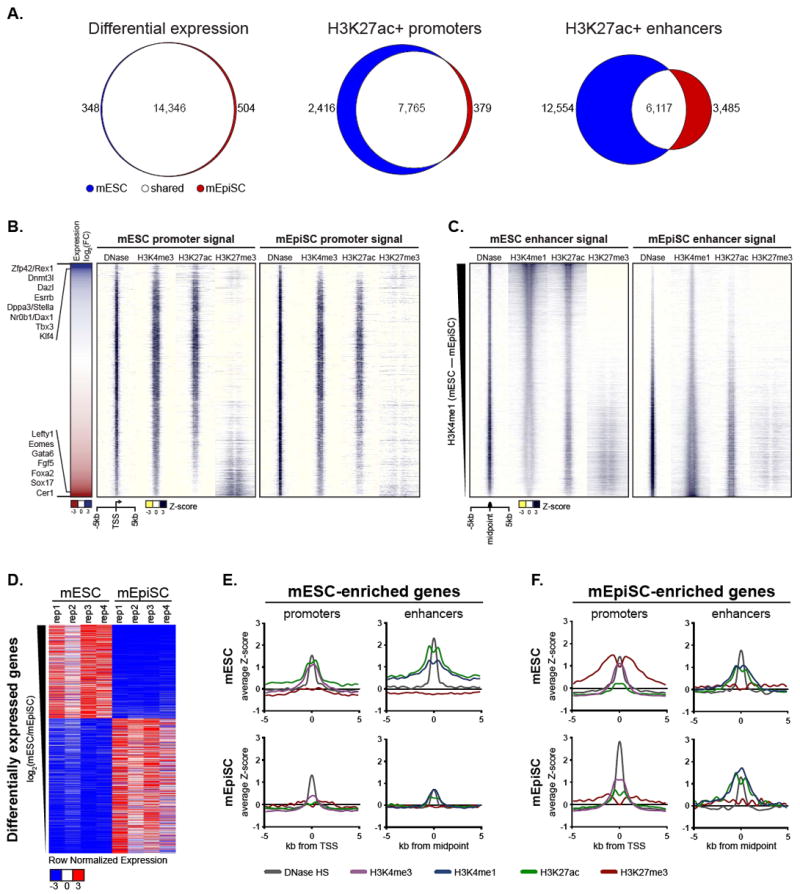
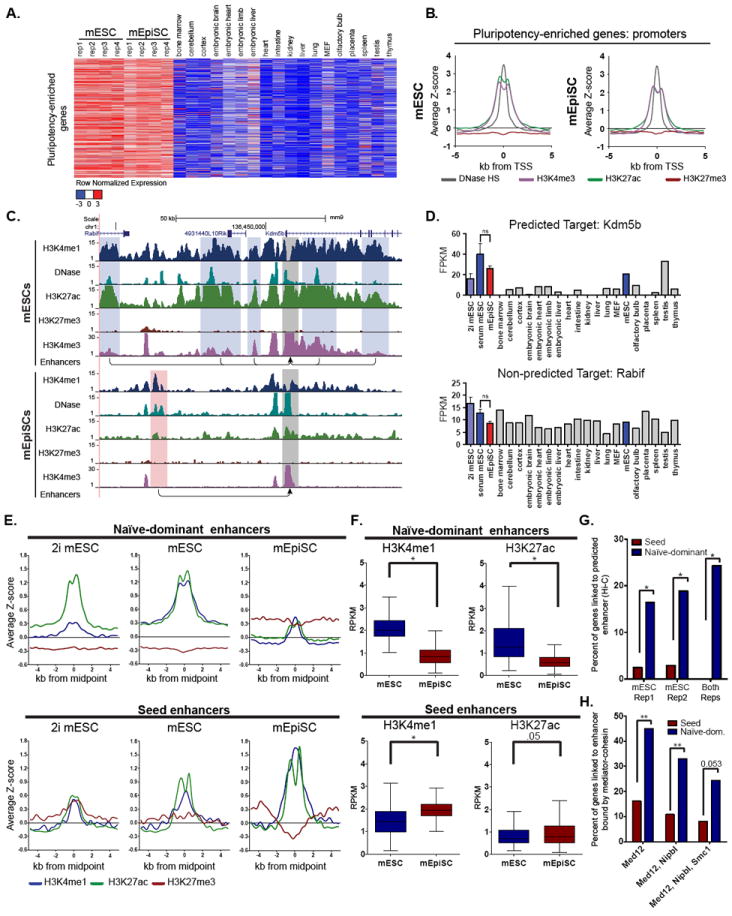
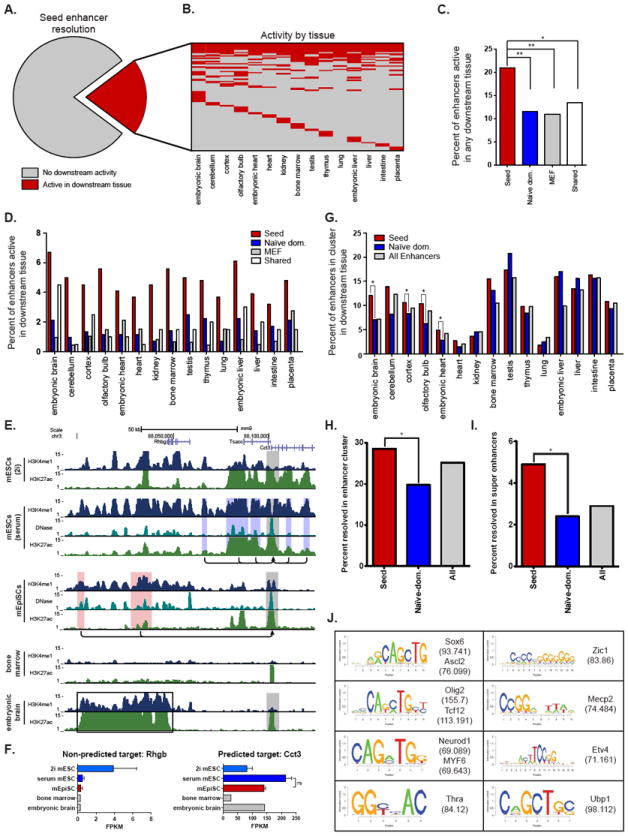
Naive mouse embryonic stem cells (mESCs) and primed epiblast stem cells (mEpiSCs) represent successive snapshots of pluripotency during embryogenesis. Using transcriptomic and epigenomic mapping we show that a small fraction of transcripts are differentially expressed between mESCs and mEpiSCs and that these genes show expected changes in chromatin at their promoters and enhancers. Unexpectedly, the cis-regulatory circuitry of genes that are expressed at identical levels between these cell states also differs dramatically. In mESCs, these genes are associated with dominant proximal enhancers and dormant distal enhancers, which we term seed enhancers. In mEpiSCs, the naive-dominant enhancers are lost, and the seed enhancers take up primary transcriptional control. Seed enhancers have increased sequence conservation and show preferential usage in downstream somatic tissues, often expanding into super enhancers. We propose that seed enhancers ensure proper enhancer utilization and transcriptional fidelity as mammalian cells transition from naive pluripotency to a somatic regulatory program.
Copyright © 2014 Elsevier Inc. All rights reserved.
Conflict of interest statement
All authors declare no competing financial interests.
Figures
References
-
- Bradley A, Evans M, Kaufman MH, Robertson E. Formation of germ-line chimaeras from embryo-derived teratocarcinoma cell lines. Nature. 1984;309:255–256. - PubMed
-
- Brons IG, Smithers LE, Trotter MW, Rugg-Gunn P, Sun B, Chuva de Sousa Lopes SM, Howlett SK, Clarkson A, Ahrlund-Richter L, Pedersen RA, et al. Derivation of pluripotent epiblast stem cells from mammalian embryos. Nature. 2007;448:191–195. - PubMed
-
- Chan YS, Goke J, Ng JH, Lu X, Gonzales KA, Tan CP, Tng WQ, Hong ZZ, Lim YS, Ng HH. Induction of a human pluripotent state with distinct regulatory circuitry that resembles preimplantation epiblast. Cell stem cell. 2013;13:663–675. - PubMed
-
- Chenoweth JG, McKay RD, Tesar PJ. Epiblast stem cells contribute new insight into pluripotency and gastrulation. Dev Growth Differ. 2010;52:293–301. - PubMed
Publication types
MeSH terms
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
