

Eosinophils and type 2 cytokine signaling in macrophages orchestrate development of functional beige fat
- PMID: 24906148
- PMCID: PMC4129510
- DOI: 10.1016/j.cell.2014.03.066
Eosinophils and type 2 cytokine signaling in macrophages orchestrate development of functional beige fat
Abstract
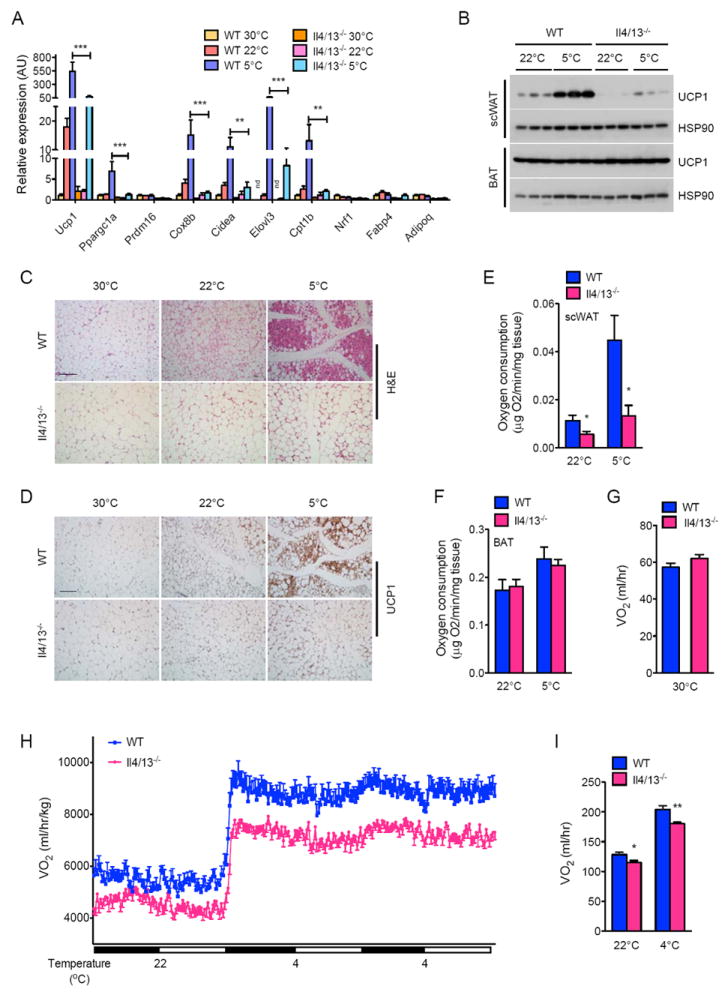
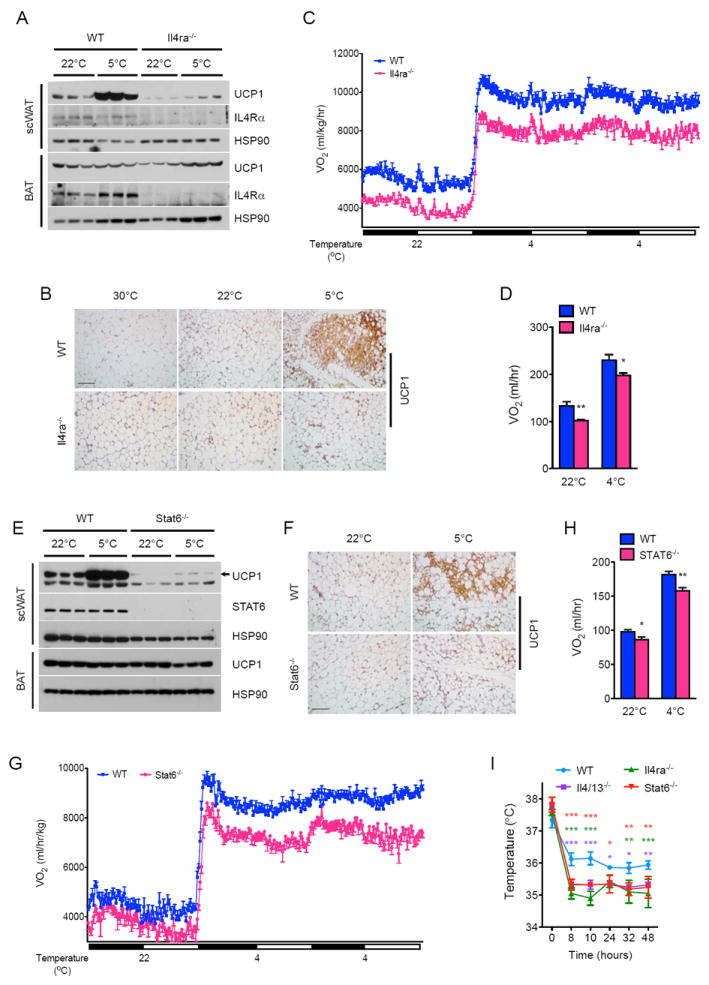
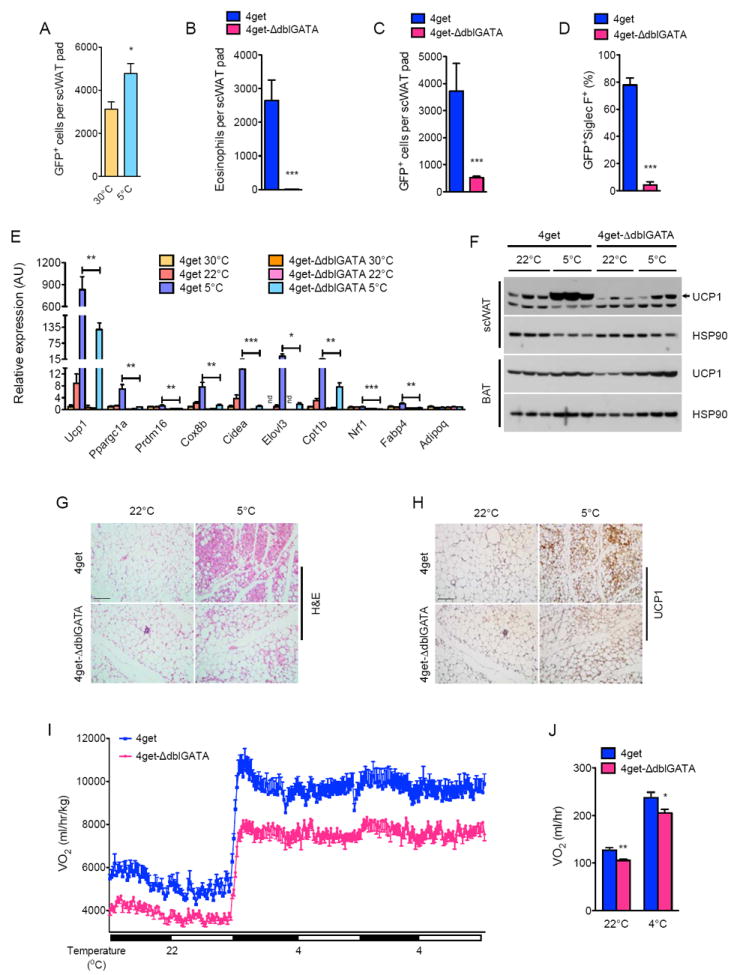
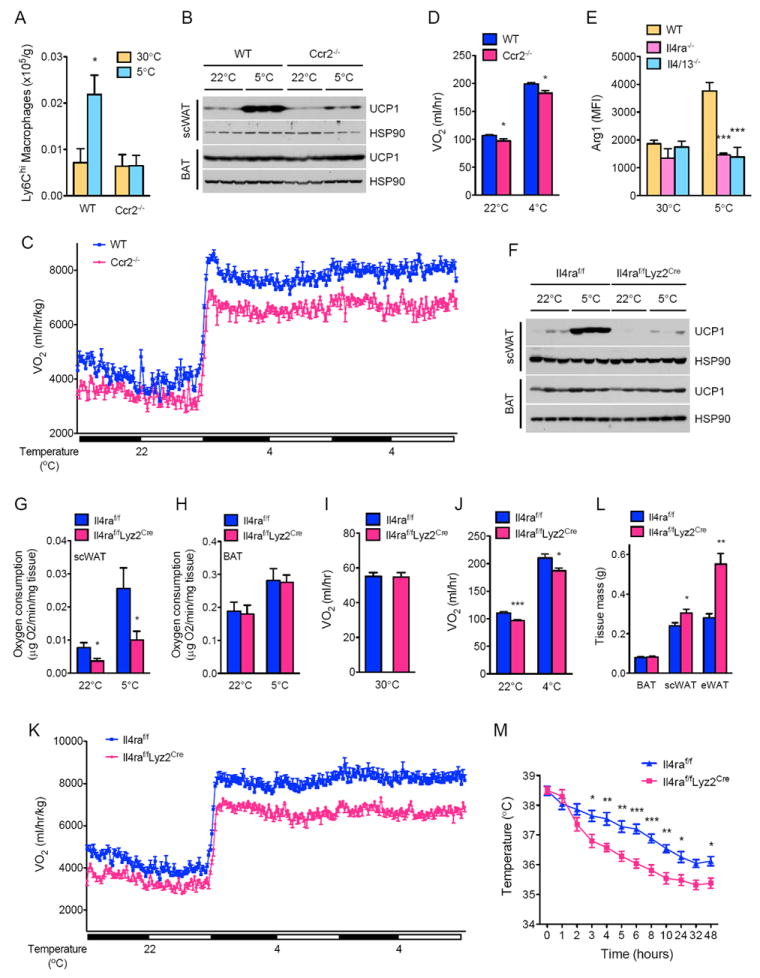
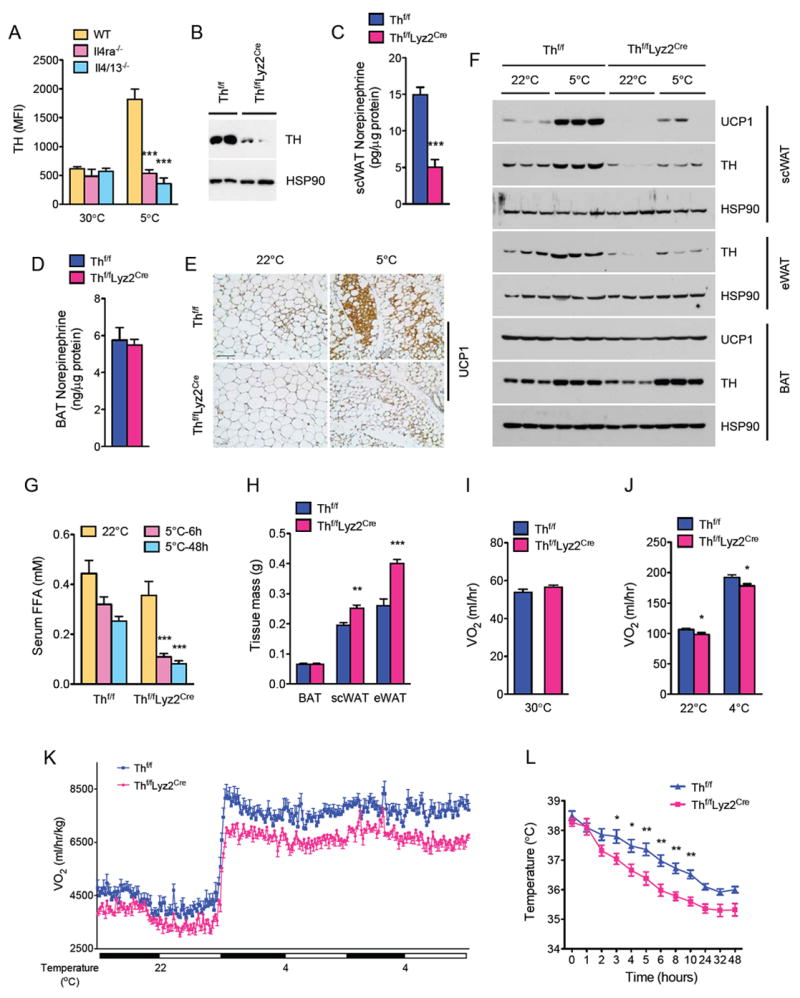
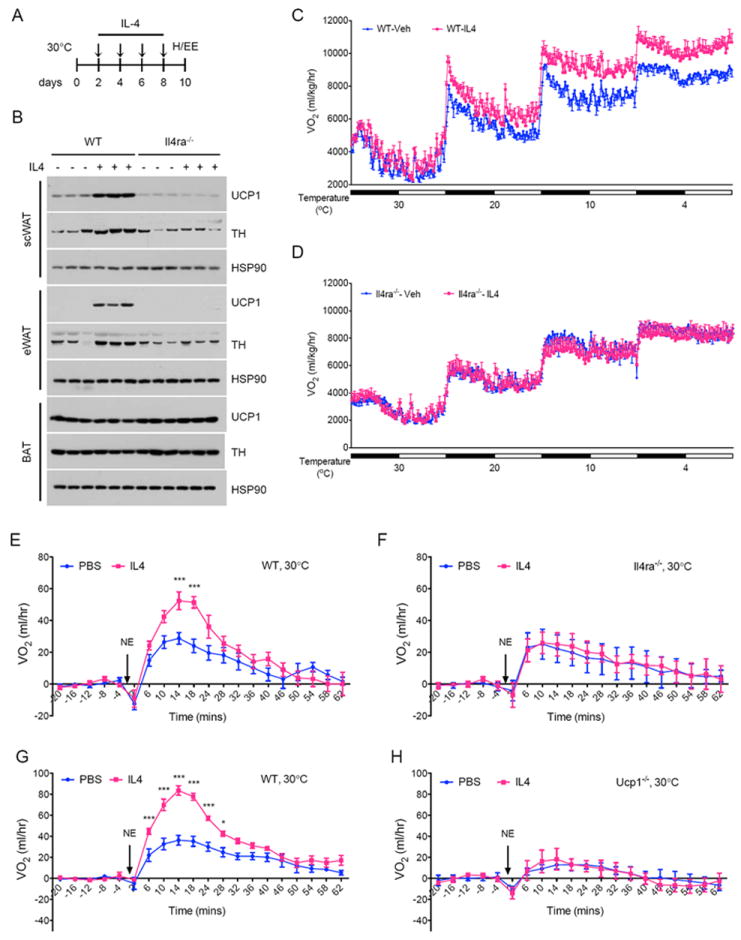
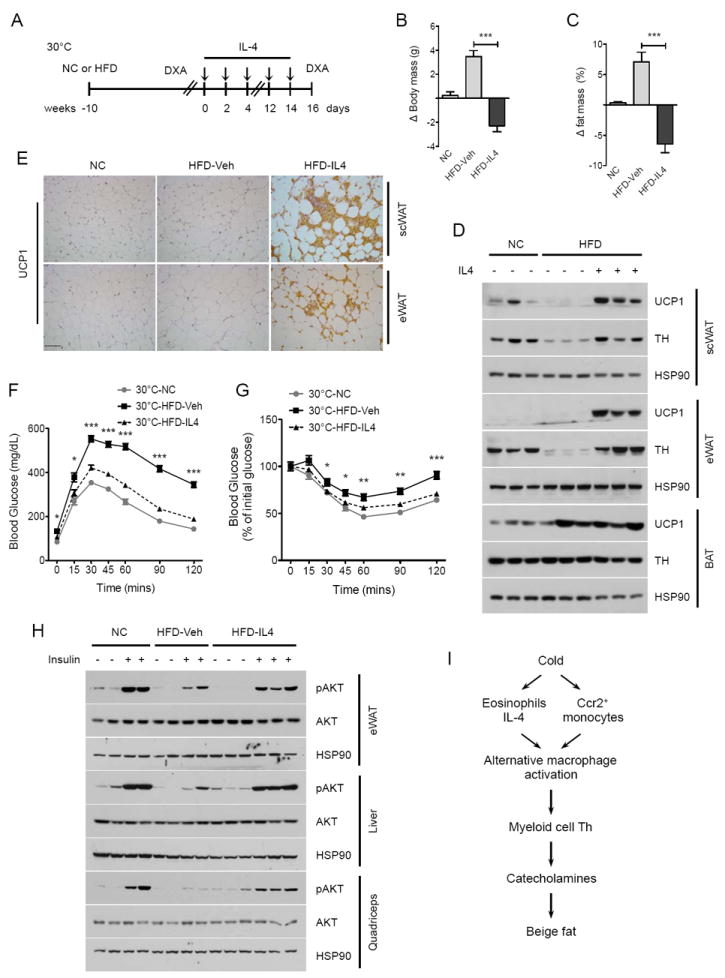
Beige fat, which expresses the thermogenic protein UCP1, provides a defense against cold and obesity. Although a cold environment is the physiologic stimulus for inducing beige fat in mice and humans, the events that lead from the sensing of cold to the development of beige fat remain poorly understood. Here, we identify the efferent beige fat thermogenic circuit, consisting of eosinophils, type 2 cytokines interleukin (IL)-4/13, and alternatively activated macrophages. Genetic loss of eosinophils or IL-4/13 signaling impairs cold-induced biogenesis of beige fat. Mechanistically, macrophages recruited to cold-stressed subcutaneous white adipose tissue (scWAT) undergo alternative activation to induce tyrosine hydroxylase expression and catecholamine production, factors required for browning of scWAT. Conversely, administration of IL-4 to thermoneutral mice increases beige fat mass and thermogenic capacity to ameliorate pre-established obesity. Together, our findings have uncovered the efferent circuit controlling biogenesis of beige fat and provide support for its targeting to treat obesity.
Copyright © 2014 Elsevier Inc. All rights reserved.
Conflict of interest statement
The authors declare that they have no competing financial interests.
Figures
Comment in
-
Eosinophils in fat: pink is the new brown.Cell. 2014 Jun 5;157(6):1249-1250. doi: 10.1016/j.cell.2014.05.025. Cell. 2014. PMID: 24906141
-
Metabolism: Type 2 immunity at the origin of beige adipocytes.Nat Rev Endocrinol. 2014 Aug;10(8):443. doi: 10.1038/nrendo.2014.99. Epub 2014 Jun 24. Nat Rev Endocrinol. 2014. PMID: 24958309 No abstract available.
References
-
- Cannon B, Nedergaard J. Metabolic consequences of the presence or absence of the thermogenic capacity of brown adipose tissue in mice (and probably in humans) Int J Obes (Lond) 2010;34(Suppl 1):S7–16. - PubMed
-
- Cannon B, Nedergaard J. Nonshivering thermogenesis and its adequate measurement in metabolic studies. The Journal of experimental biology. 2011;214:242–253. - PubMed
-
- Charo IF, Ransohoff RM. The many roles of chemokines and chemokine receptors in inflammation. The New England journal of medicine. 2006;354:610–621. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
