

Molecular basis for age-dependent microtubule acetylation by tubulin acetyltransferase
- PMID: 24906155
- PMCID: PMC4726456
- DOI: 10.1016/j.cell.2014.03.061
Molecular basis for age-dependent microtubule acetylation by tubulin acetyltransferase
Abstract
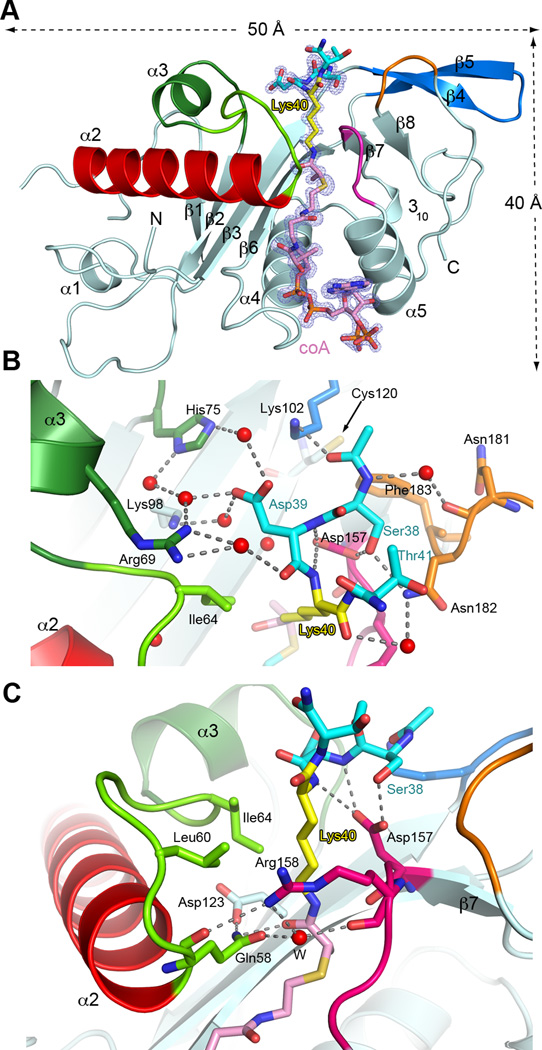
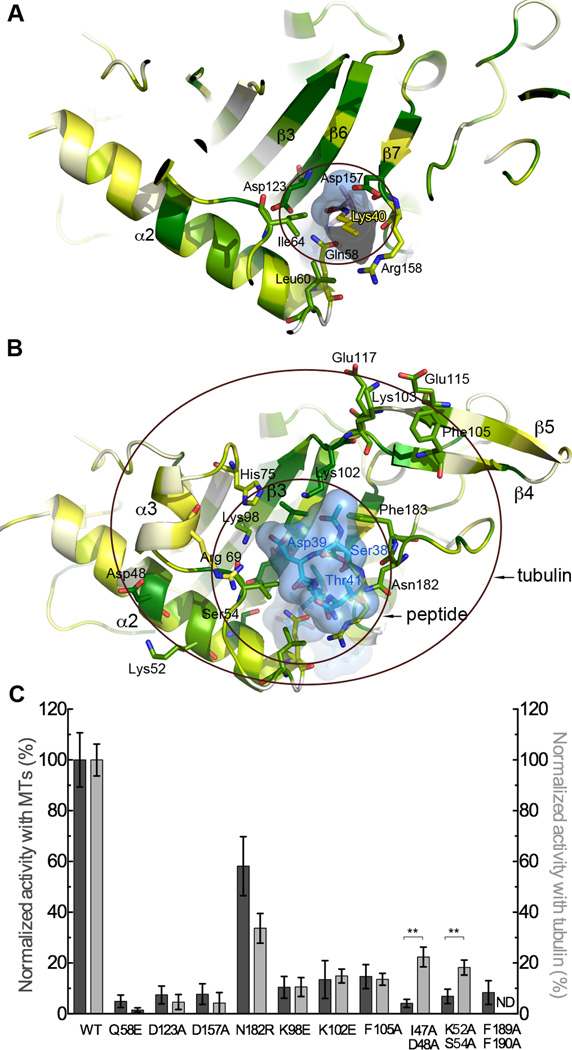
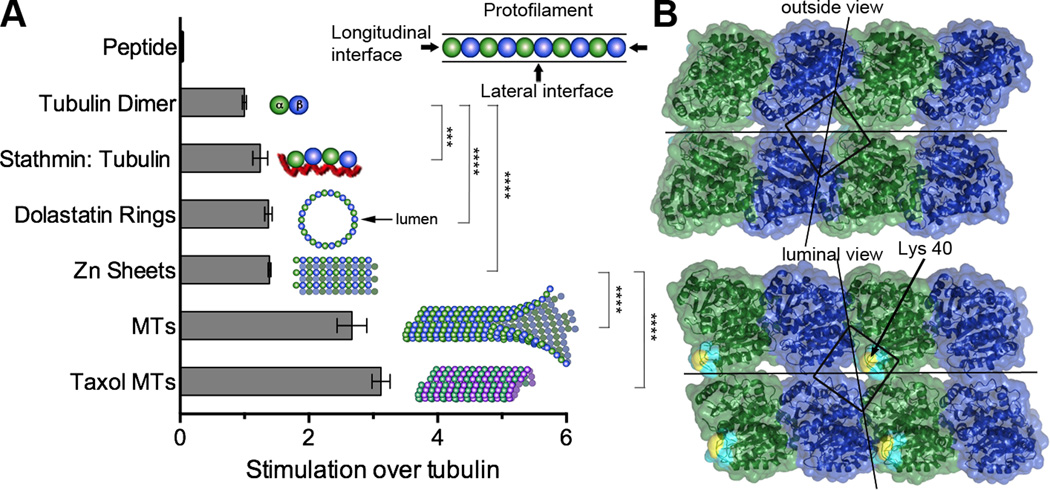
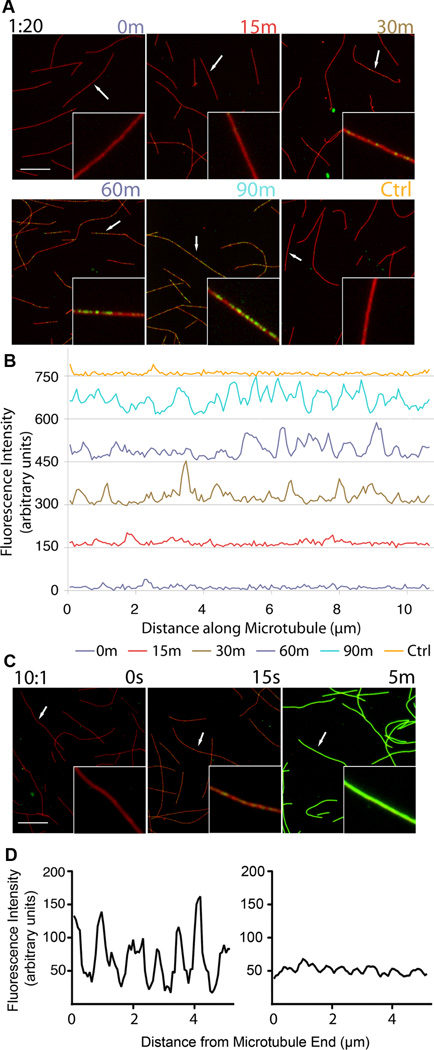
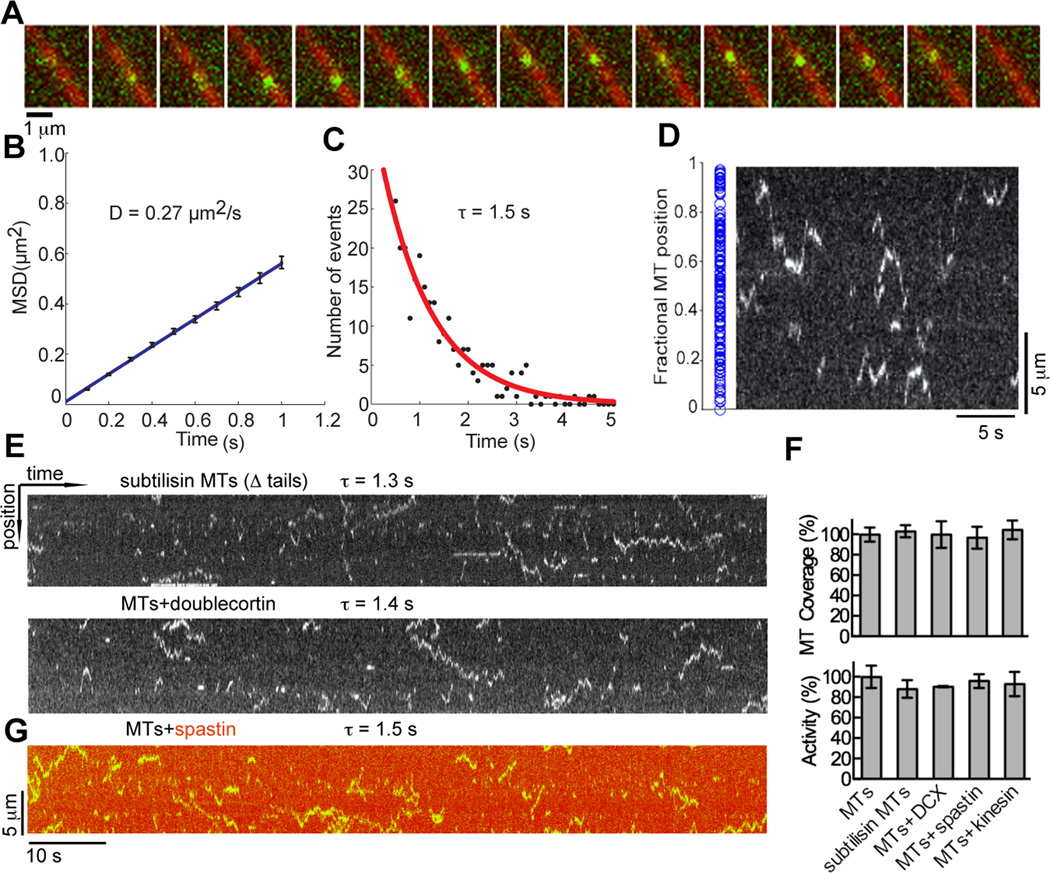
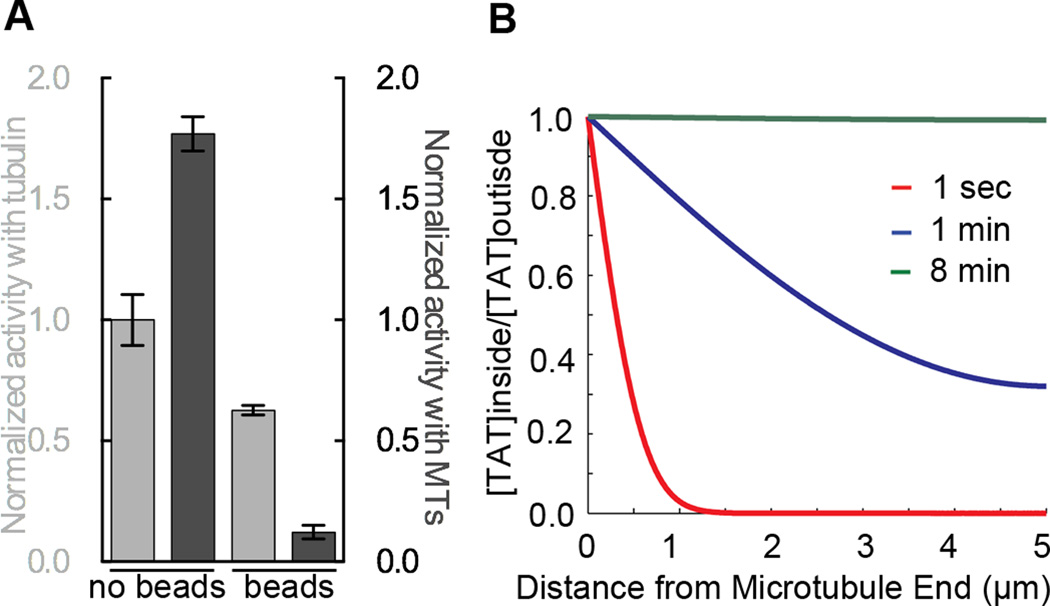
Acetylation of α-tubulin Lys40 by tubulin acetyltransferase (TAT) is the only known posttranslational modification in the microtubule lumen. It marks stable microtubules and is required for polarity establishment and directional migration. Here, we elucidate the mechanistic underpinnings for TAT activity and its preference for microtubules with slow turnover. 1.35 Å TAT cocrystal structures with bisubstrate analogs constrain TAT action to the microtubule lumen and reveal Lys40 engaged in a suboptimal active site. Assays with diverse tubulin polymers show that TAT is stimulated by microtubule interprotofilament contacts. Unexpectedly, despite the confined intraluminal location of Lys40, TAT efficiently scans the microtubule bidirectionally and acetylates stochastically without preference for ends. First-principles modeling and single-molecule measurements demonstrate that TAT catalytic activity, not constrained luminal diffusion, is rate limiting for acetylation. Thus, because of its preference for microtubules over free tubulin and its modest catalytic rate, TAT can function as a slow clock for microtubule lifetimes.
Copyright © 2014 Elsevier Inc. All rights reserved.
Figures
Comment in
-
A slow dance for microtubule acetylation.Cell. 2014 Jun 5;157(6):1255-1256. doi: 10.1016/j.cell.2014.05.021. Cell. 2014. PMID: 24906144
References
-
- Bungay PM, Brenner H. The motion of a closely-fitting sphere in a fluid-filled tube. Internaltional Jounral of Multiphase Flow. 1973;1:25–56.
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
