

The histone acetyltransferase GcnE (GCN5) plays a central role in the regulation of Aspergillus asexual development
- PMID: 24907261
- PMCID: PMC4125392
- DOI: 10.1534/genetics.114.165688
The histone acetyltransferase GcnE (GCN5) plays a central role in the regulation of Aspergillus asexual development
Abstract
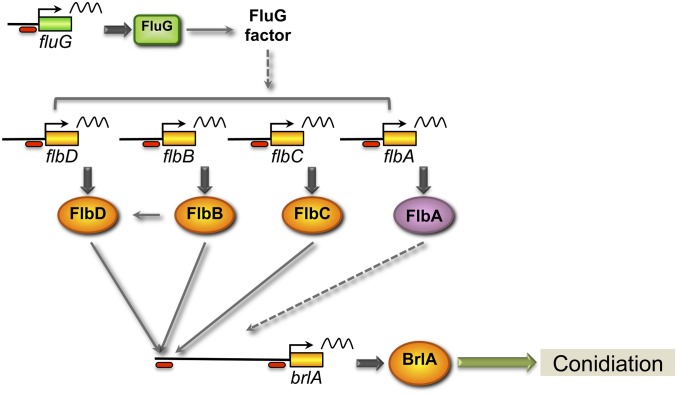
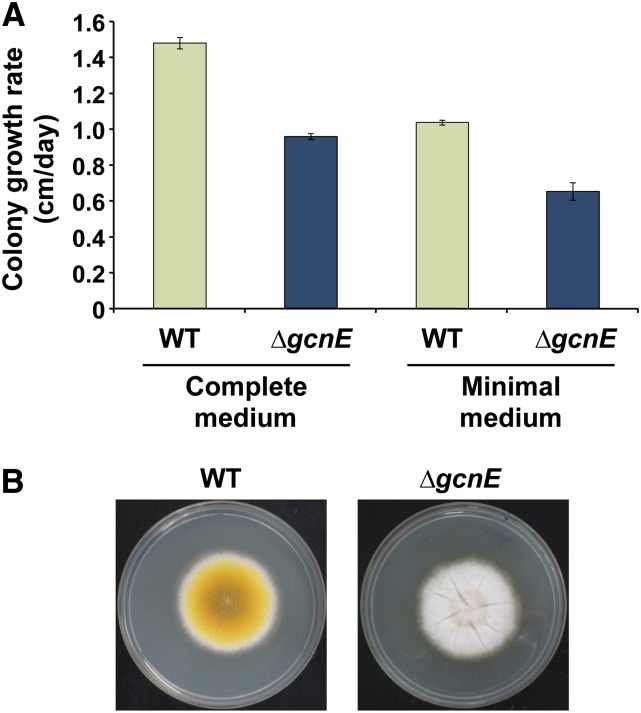
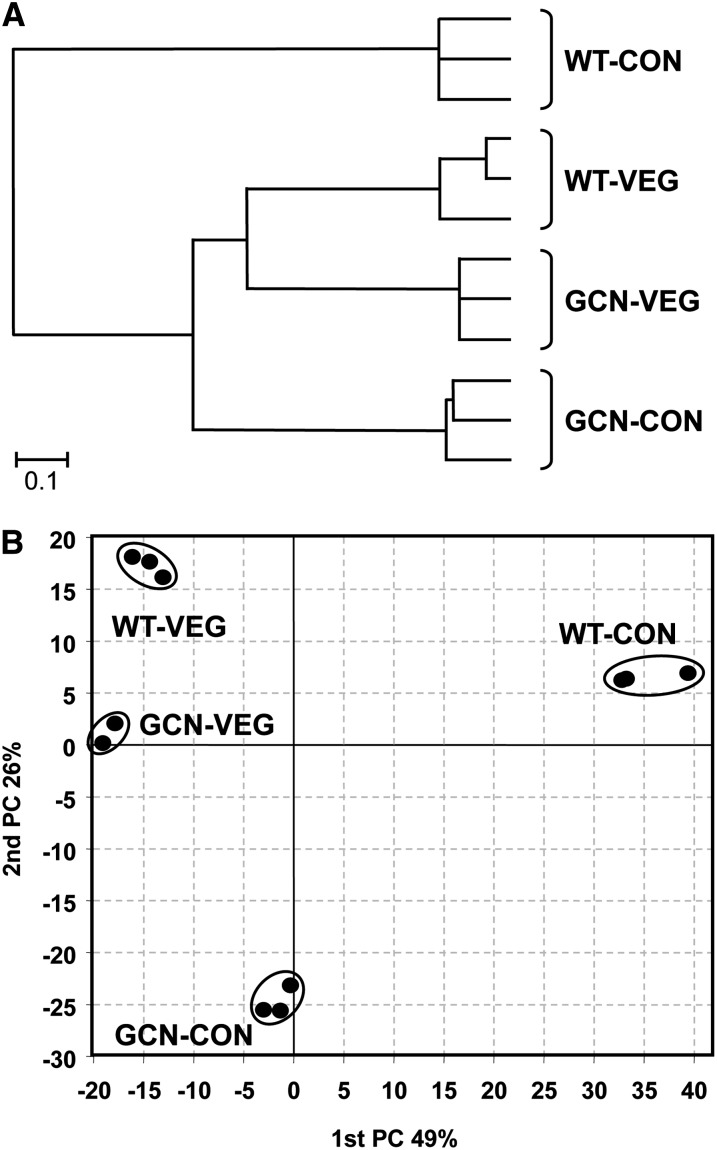
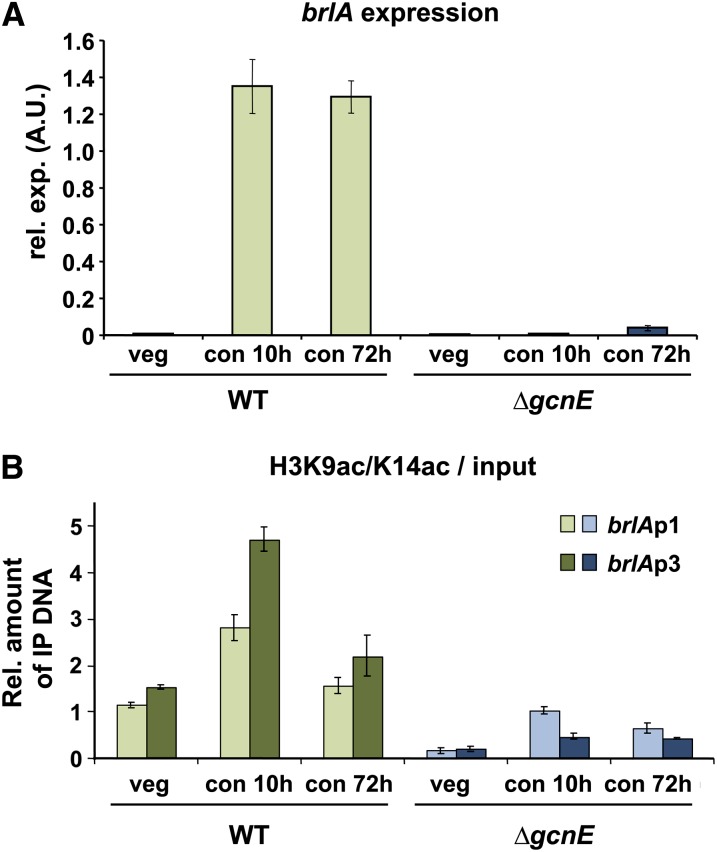
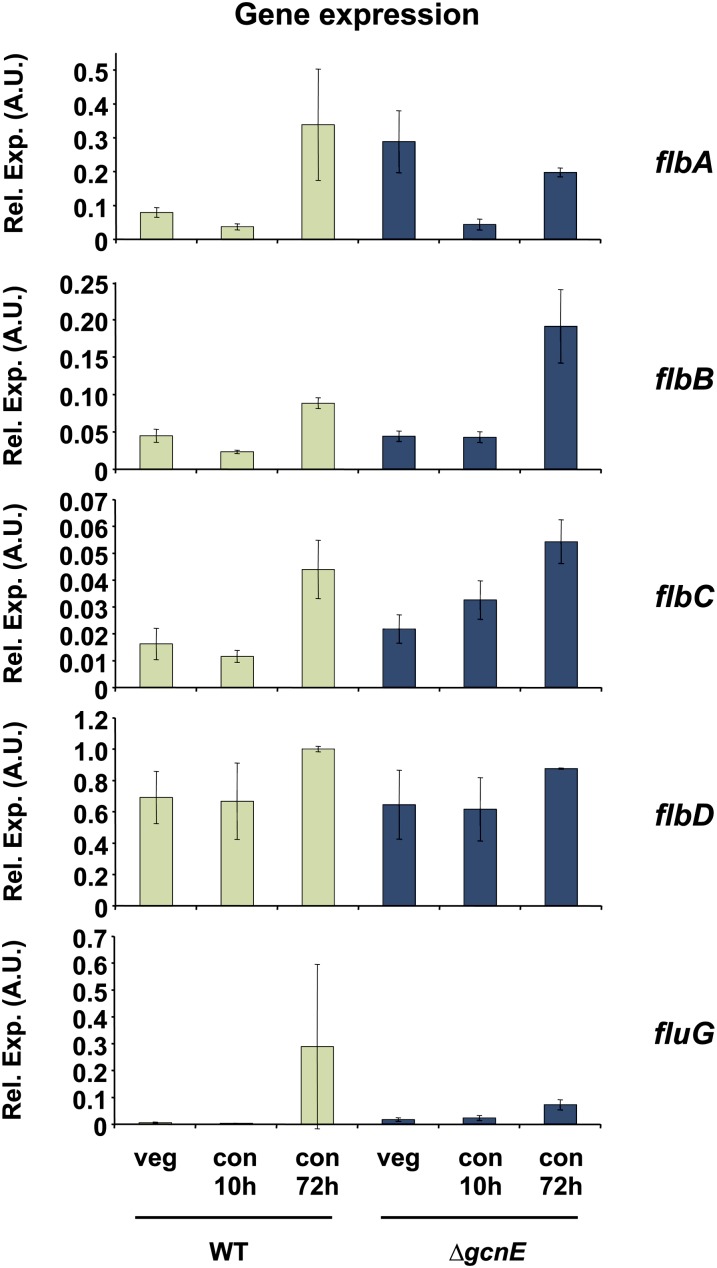
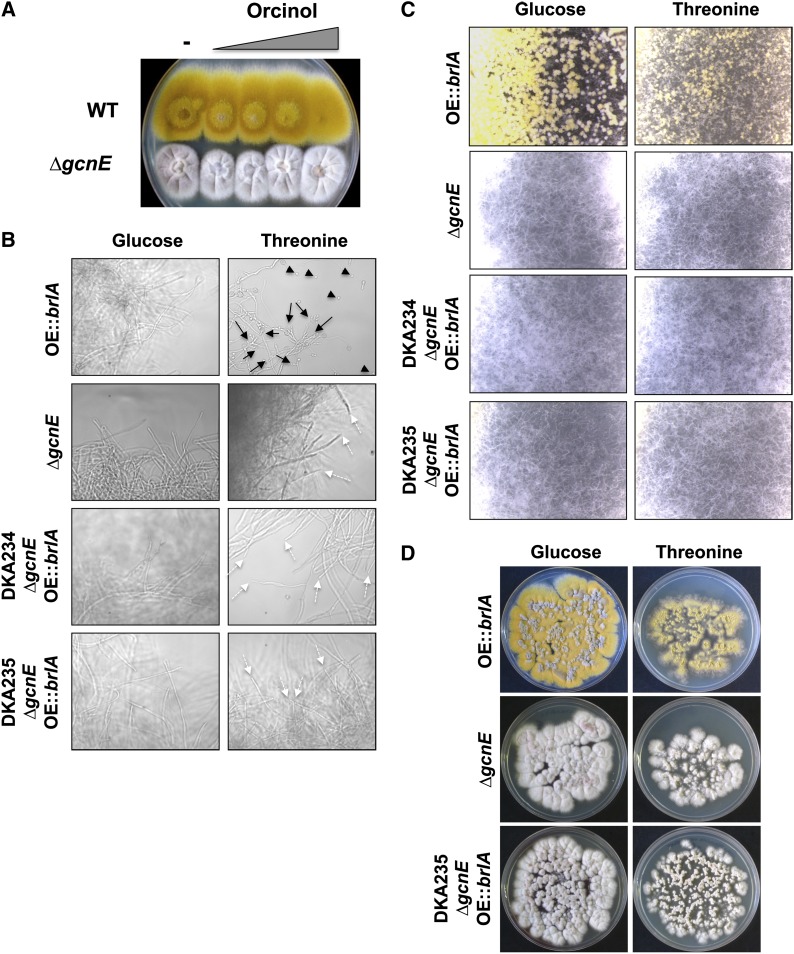
Acetylation of histones is a key regulatory mechanism of gene expression in eukaryotes. GcnE is an acetyltransferase of Aspergillus nidulans involved in the acetylation of histone H3 at lysine 9 and lysine 14. Previous works have demonstrated that deletion of gcnE results in defects in primary and secondary metabolism. Here we unveil the role of GcnE in development and show that a ∆gcnE mutant strain has minor growth defects but is impaired in normal conidiophore development. No signs of conidiation were found after 3 days of incubation, and immature and aberrant conidiophores were found after 1 week of incubation. Centroid linkage clustering and principal component (PC) analysis of transcriptomic data suggest that GcnE occupies a central position in Aspergillus developmental regulation and that it is essential for inducing conidiation genes. GcnE function was found to be required for the acetylation of histone H3K9/K14 at the promoter of the master regulator of conidiation, brlA, as well as at the promoters of the upstream developmental regulators of conidiation flbA, flbB, flbC, and flbD (fluffy genes). However, analysis of the gene expression of brlA and the fluffy genes revealed that the lack of conidiation originated in a complete absence of brlA expression in the ∆gcnE strain. Ectopic induction of brlA from a heterologous alcA promoter did not remediate the conidiation defects in the ∆gcnE strain, suggesting that additional GcnE-mediated mechanisms must operate. Therefore, we conclude that GcnE is the only nonessential histone modifier with a strong role in fungal development found so far.
Keywords: Aspergillus; Gcn5; GcnE; SAGA; asexual development; brlA; conidiation; fluffy genes; histone acetylation.
Copyright © 2014 by the Genetics Society of America.
Figures

References
-
- Adams T. H., Boylan M. T., Timberlake W. E., 1988. brlA is necessary and sufficient to direct conidiophore development in Aspergillus nidulans. Cell 54: 353–362. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
