

Retrospective on the all-in-one retroviral nucleocapsid protein
- PMID: 24907482
- PMCID: PMC7114435
- DOI: 10.1016/j.virusres.2014.05.011
Retrospective on the all-in-one retroviral nucleocapsid protein
Abstract
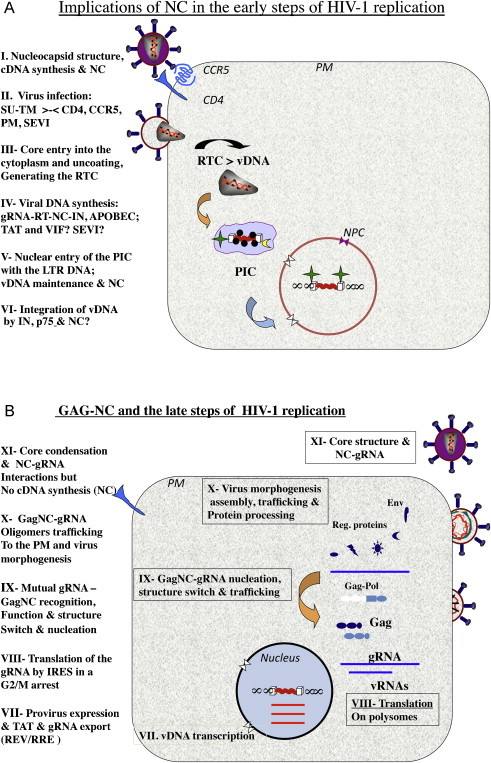
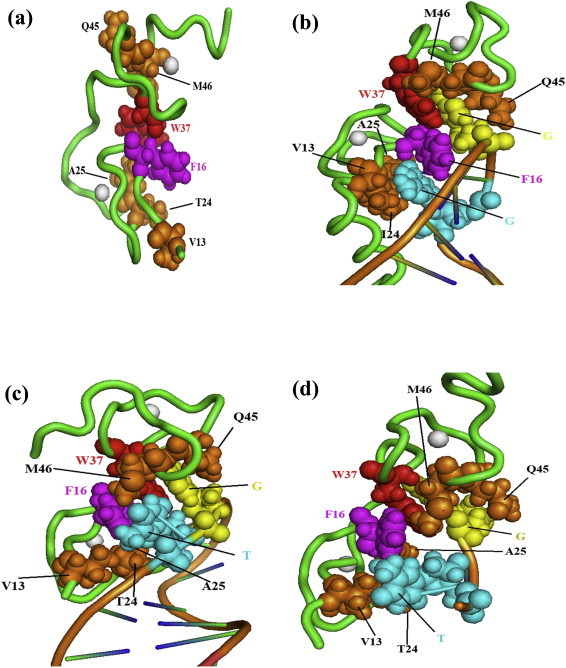
This review aims at briefly presenting a retrospect on the retroviral nucleocapsid protein (NC), from an unspecific nucleic acid binding protein (NABP) to an all-in-one viral protein with multiple key functions in the early and late phases of the retrovirus replication cycle, notably reverse transcription of the genomic RNA and viral DNA integration into the host genome, and selection of the genomic RNA together with the initial steps of virus morphogenesis. In this context we will discuss the notion that NC protein has a flexible conformation and is thus a member of the growing family of intrinsically disordered proteins (IDPs) where disorder may account, at least in part, for its function as a nucleic acid (NA) chaperone and possibly as a protein chaperone vis-à-vis the viral DNA polymerase during reverse transcription. Lastly, we will briefly review the development of new anti-retroviral/AIDS compounds targeting HIV-1 NC because it represents an ideal target due to its multiple roles in the early and late phases of virus replication and its high degree of conservation.
Keywords: Intrinsically disordered protein; Nucleocapsid protein; Retrovirus; Viral DNA synthesis; Virus formation; Zinc fingers.
Copyright © 2014 Elsevier B.V. All rights reserved.
Figures
References
-
- Abbink T.E.M., Berkhout B. A novel long distance base-pairing interaction in human immunodeficiency virus type 1 RNA occludes the Gag start codon. J. Biol. Chem. 2003;278:11601–11611. - PubMed
-
- Adamson C.S., Freed E.O. Human immunodeficiency virus type 1 assembly, release, and maturation. Adv. Pharmacol. (San Diego, CA) 2007;55:347–387. - PubMed
-
- Allain B., Rascle J.B., de Rocquigny H., Roques B., Darlix J.L. CIS elements and trans-acting factors required for minus strand DNA transfer during reverse transcription of the genomic RNA of murine leukemia virus. J. Mol. Biol. 1998;277:225–235. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
