

Quantitative genome-wide enhancer activity maps for five Drosophila species show functional enhancer conservation and turnover during cis-regulatory evolution
- PMID: 24908250
- PMCID: PMC4250274
- DOI: 10.1038/ng.3009
Quantitative genome-wide enhancer activity maps for five Drosophila species show functional enhancer conservation and turnover during cis-regulatory evolution
Abstract
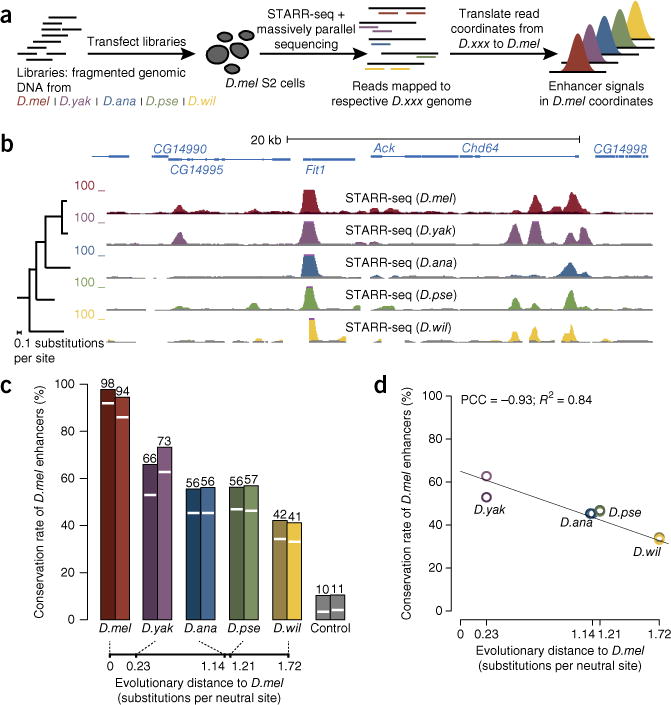
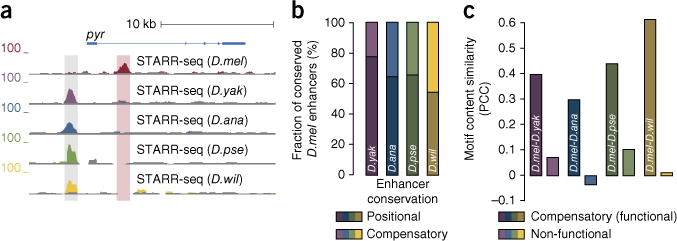
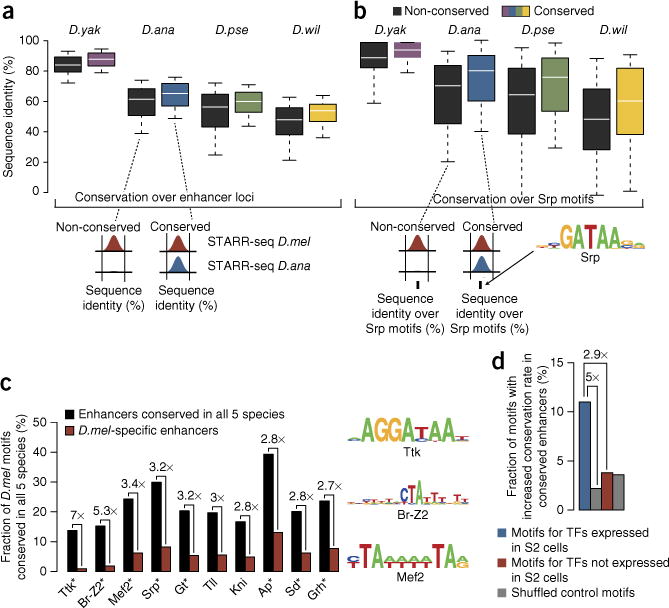
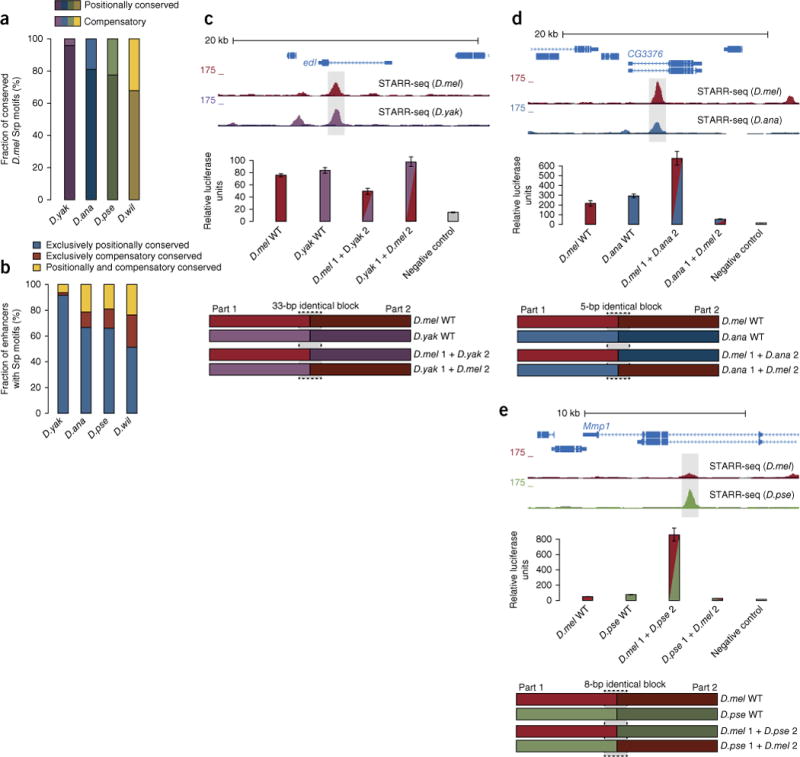
Phenotypic differences between closely related species are thought to arise primarily from changes in gene expression due to mutations in cis-regulatory sequences (enhancers). However, it has remained unclear how frequently mutations alter enhancer activity or create functional enhancers de novo. Here we use STARR-seq, a recently developed quantitative enhancer assay, to determine genome-wide enhancer activity profiles for five Drosophila species in the constant trans-regulatory environment of Drosophila melanogaster S2 cells. We find that the functions of a large fraction of D. melanogaster enhancers are conserved for their orthologous sequences owing to selection and stabilizing turnover of transcription factor motifs. Moreover, hundreds of enhancers have been gained since the D. melanogaster-Drosophila yakuba split about 11 million years ago without apparent adaptive selection and can contribute to changes in gene expression in vivo. Our finding that enhancer activity is often deeply conserved and frequently gained provides functional insights into regulatory evolution.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
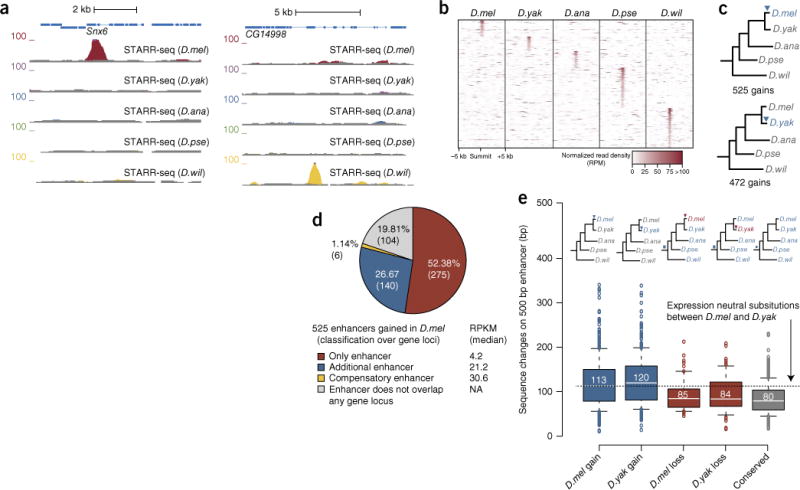
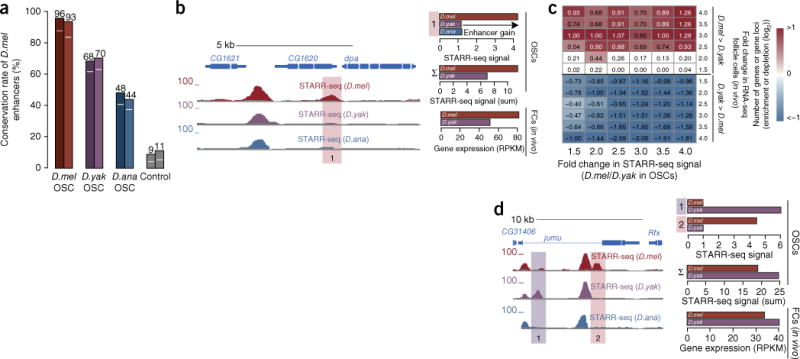
References
-
- Carroll SB. Evo-devo and an expanding evolutionary synthesis: a genetic theory of morphological evolution. Cell. 2008;134:25–36. - PubMed
-
- King MC, Wilson AC. Evolution at two levels in humans and chimpanzees. Science. 1975;188:107–116. - PubMed
-
- Wittkopp PJ, Kalay G. Cis-regulatory elements: molecular mechanisms and evolutionary processes underlying divergence. Nat Rev Genet. 2012;13:59–69. - PubMed
-
- Wray GA. The evolutionary significance of cis-regulatory mutations. Nat Rev Genet. 2007;8:206–216. - PubMed
-
- Gompel N, Prud’homme B, Wittkopp PJ, Kassner VA, Carroll SB. Chance caught on the wing: cis-regulatory evolution and the origin of pigment patterns in Drosophila. Nature. 2005;433:481–487. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
