

Amygdala microcircuits controlling learned fear
- PMID: 24908482
- PMCID: PMC4103014
- DOI: 10.1016/j.neuron.2014.04.042
Amygdala microcircuits controlling learned fear
Abstract
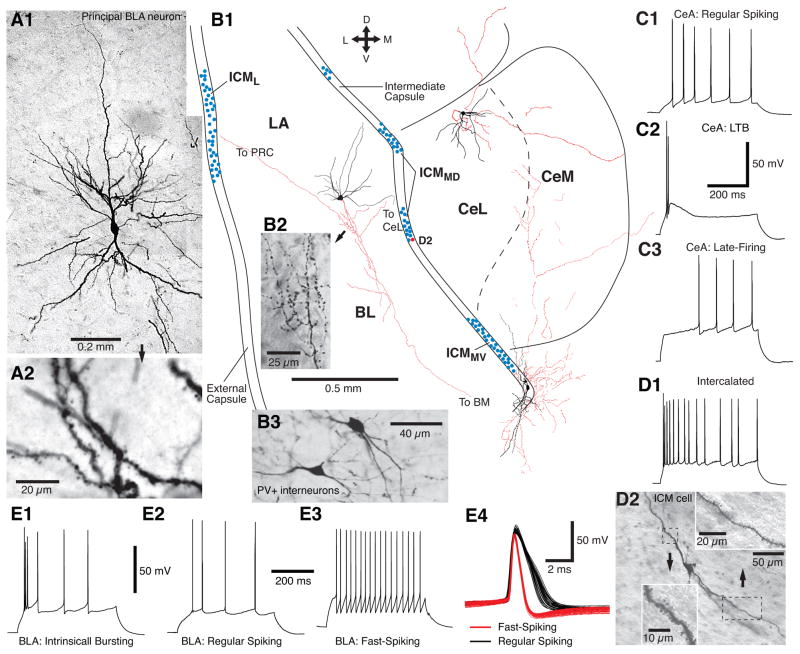
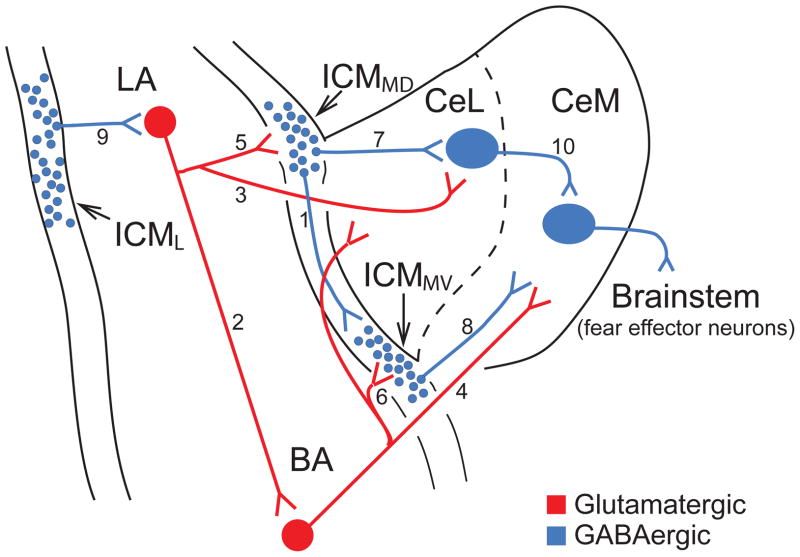
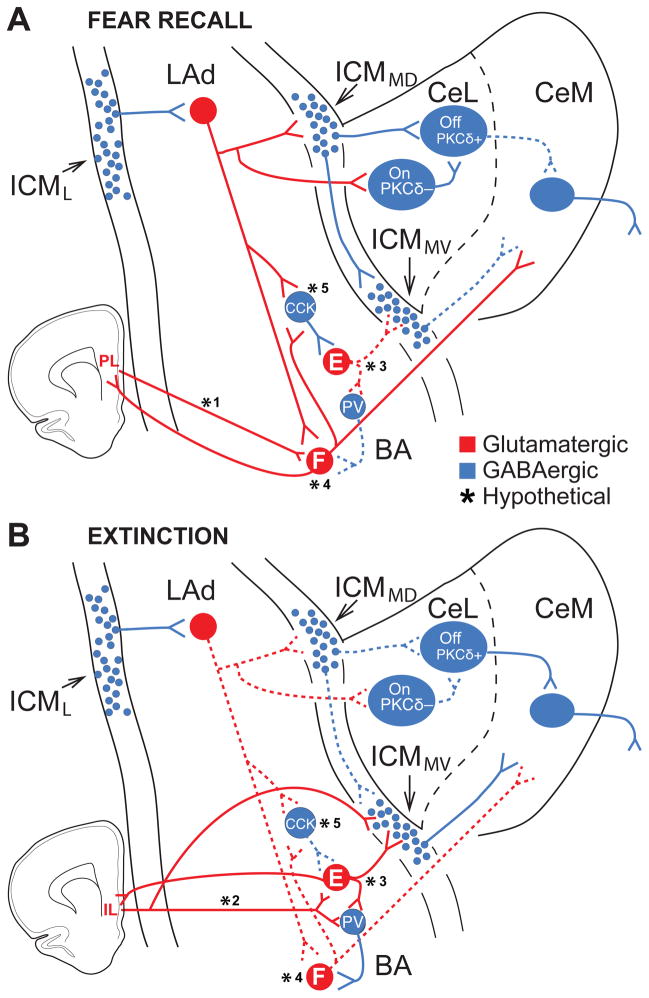
We review recent work on the role of intrinsic amygdala networks in the regulation of classically conditioned defensive behaviors, commonly known as conditioned fear. These new developments highlight how conditioned fear depends on far more complex networks than initially envisioned. Indeed, multiple parallel inhibitory and excitatory circuits are differentially recruited during the expression versus extinction of conditioned fear. Moreover, shifts between expression and extinction circuits involve coordinated interactions with different regions of the medial prefrontal cortex. However, key areas of uncertainty remain, particularly with respect to the connectivity of the different cell types. Filling these gaps in our knowledge is important because much evidence indicates that human anxiety disorders results from an abnormal regulation of the networks supporting fear learning.
Copyright © 2014 Elsevier Inc. All rights reserved.
Figures
References
-
- Amorapanth P, LeDoux JE, Nader K. Different lateral amygdala outputs mediate reactions and actions elicited by a fear-arousing stimulus. Nat Neurosci. 2000;3:74–79. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
