

Age-related homeostatic midchannel proteolysis of neuronal L-type voltage-gated Ca²⁺ channels
- PMID: 24908485
- PMCID: PMC4052215
- DOI: 10.1016/j.neuron.2014.04.017
Age-related homeostatic midchannel proteolysis of neuronal L-type voltage-gated Ca²⁺ channels
Abstract
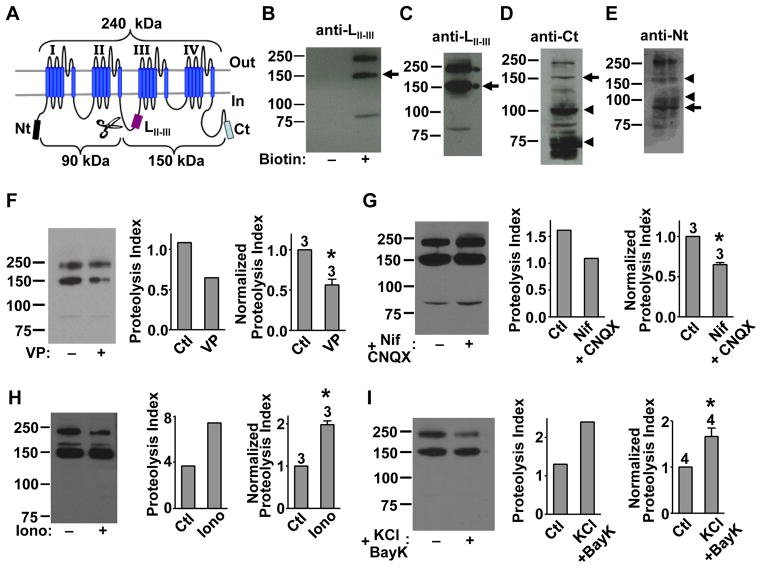
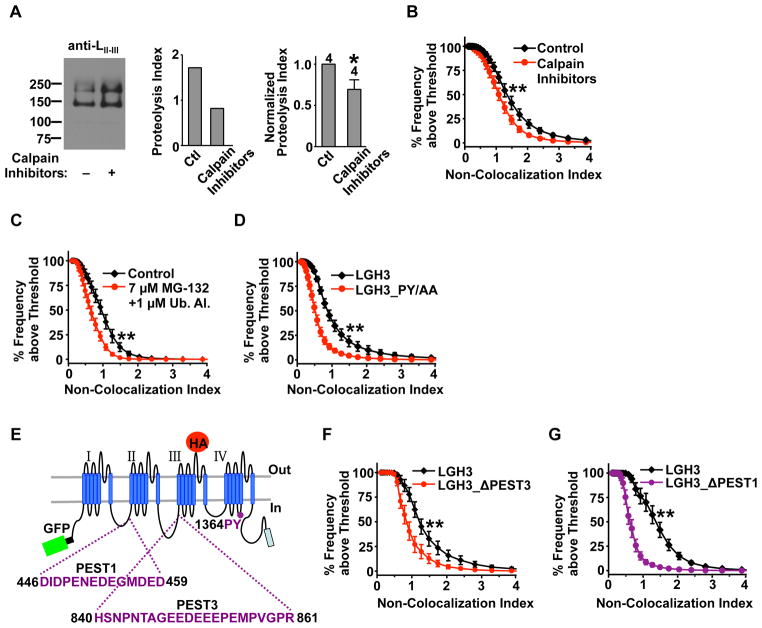
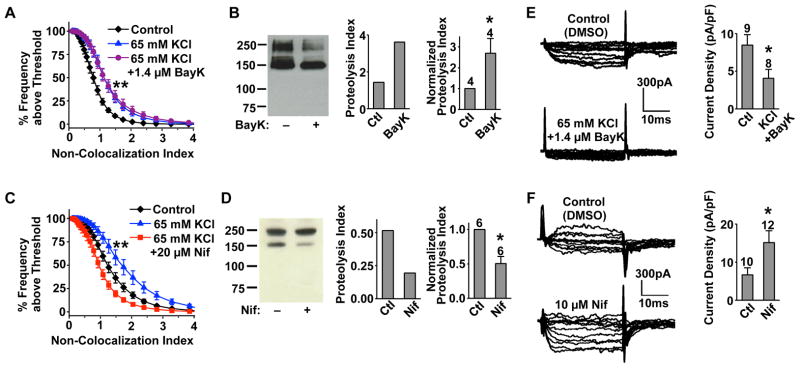
Neural circuitry and brain activity depend critically on proper function of voltage-gated calcium channels (VGCCs), whose activity must be tightly controlled. We show that the main body of the pore-forming α1 subunit of neuronal L-type VGCCs, Cav1.2, is proteolytically cleaved, resulting in Cav1.2 fragment channels that separate but remain on the plasma membrane. This "midchannel" proteolysis is regulated by channel activity, involves the Ca(2+)-dependent protease calpain and the ubiquitin-proteasome system, and causes attenuation and biophysical alterations of VGCC currents. Recombinant Cav1.2 fragment channels mimicking the products of midchannel proteolysis do not form active channels on their own but, when properly paired, produce currents with distinct biophysical properties. Midchannel proteolysis increases dramatically with age and can be attenuated with an L-type VGCC blocker in vivo. Midchannel proteolysis represents a novel form of homeostatic negative-feedback processing of VGCCs that could profoundly affect neuronal excitability, neurotransmission, neuroprotection, and calcium signaling in physiological and disease states.
Copyright © 2014 Elsevier Inc. All rights reserved.
Figures




Comment in
-
CaV1.2 calcium channels: just cut out to be regulated?Neuron. 2014 Jun 4;82(5):939-40. doi: 10.1016/j.neuron.2014.05.030. Neuron. 2014. PMID: 24908477 Free PMC article.
References
-
- Altier C, Garcia-Caballero A, Simms B, You H, Chen L, Walcher J, Tedford HW, Hermosilla T, Zamponi GW. The Cavβ subunit prevents RFP2-mediated ubiquitination and proteasomal degradation of L-type channels. Nat Neurosci. 2011;14:173–180. - PubMed
-
- Blanpied TA, Scott DB, Ehlers MD. Dynamics and regulation of clathrin coats at specialized endocytic zones of dendrites and spines. Neuron. 2002;36:435–449. - PubMed
-
- Brawley RM, Hosey MM. Identification of two distinct proteins that are immunologically related to the α1 subunit of the skeletal muscle dihydropyridine-sensitive calcium channel. J Biol Chem. 1992;267:18218–18223. - PubMed
-
- Cain SM, Snutch TP. Voltage-gated calcium channels and disease. Biofactors. 2011;37:197–205. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
