

Differential developmental requirement and peripheral regulation for dermal Vγ4 and Vγ6T17 cells in health and inflammation
- PMID: 24909159
- PMCID: PMC4068267
- DOI: 10.1038/ncomms4986
Differential developmental requirement and peripheral regulation for dermal Vγ4 and Vγ6T17 cells in health and inflammation
Erratum in
-
Corrigendum: Differential developmental requirement and peripheral regulation for dermal Vγ4 and Vγ6T17 cells in health and inflammation.Nat Commun. 2016 Apr 11;7:11354. doi: 10.1038/ncomms11354. Nat Commun. 2016. PMID: 27063645 Free PMC article. No abstract available.
Abstract
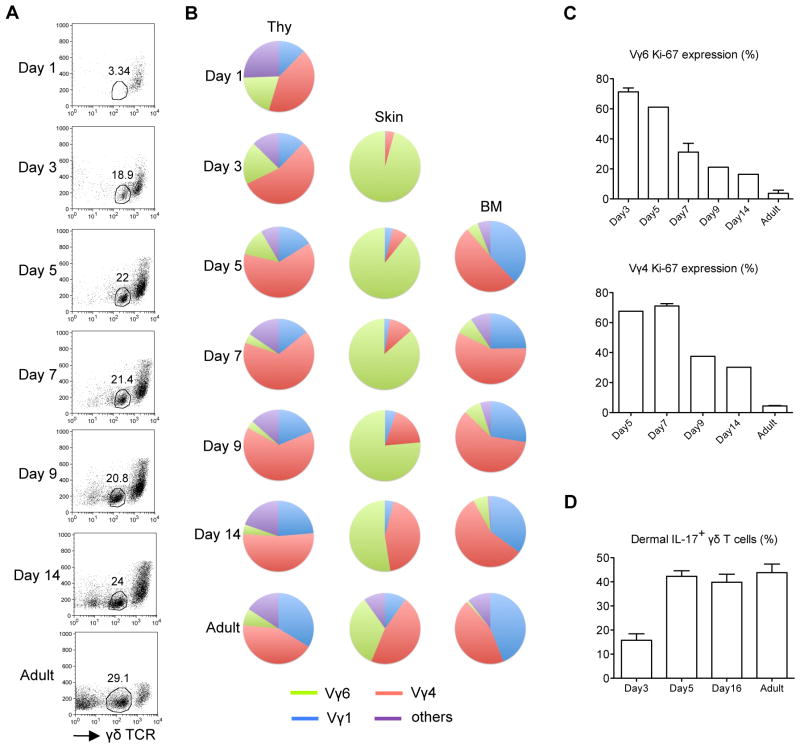
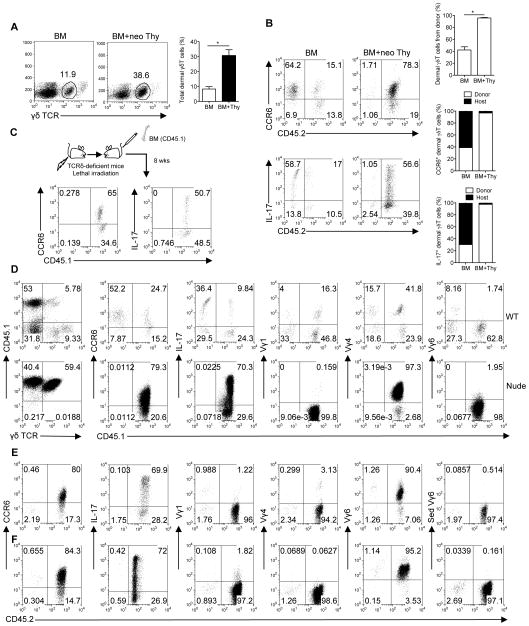
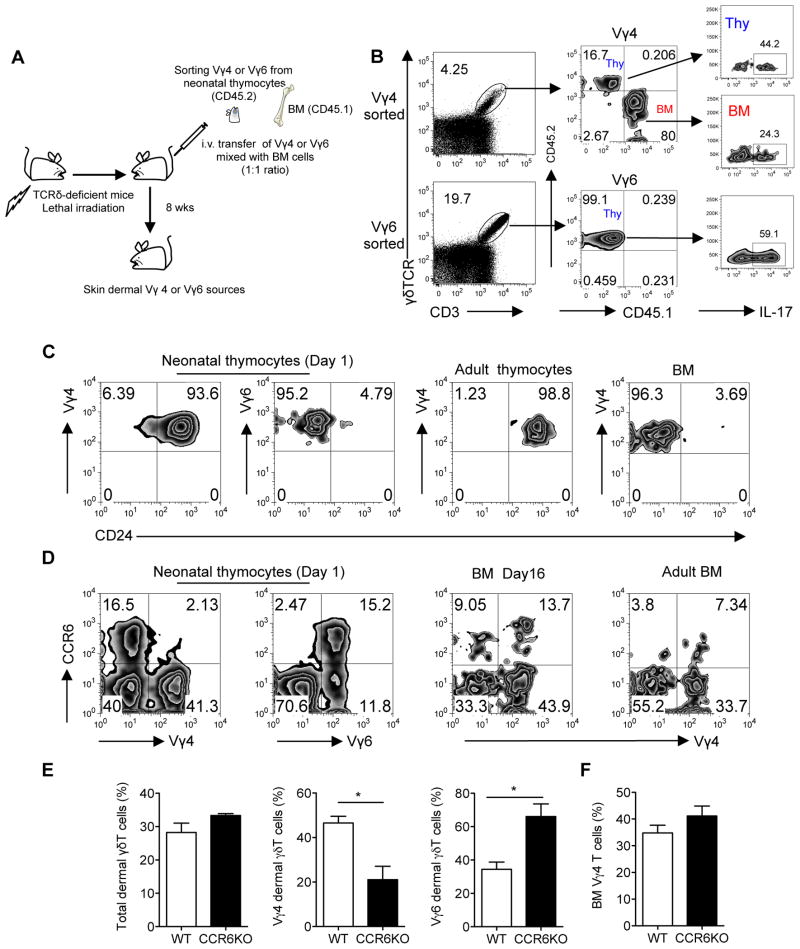
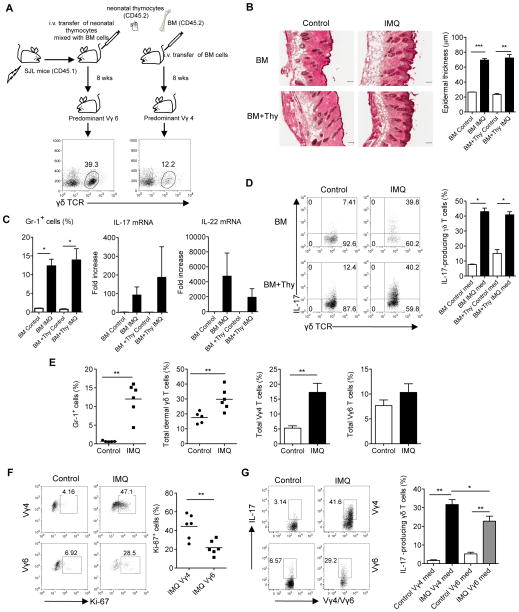
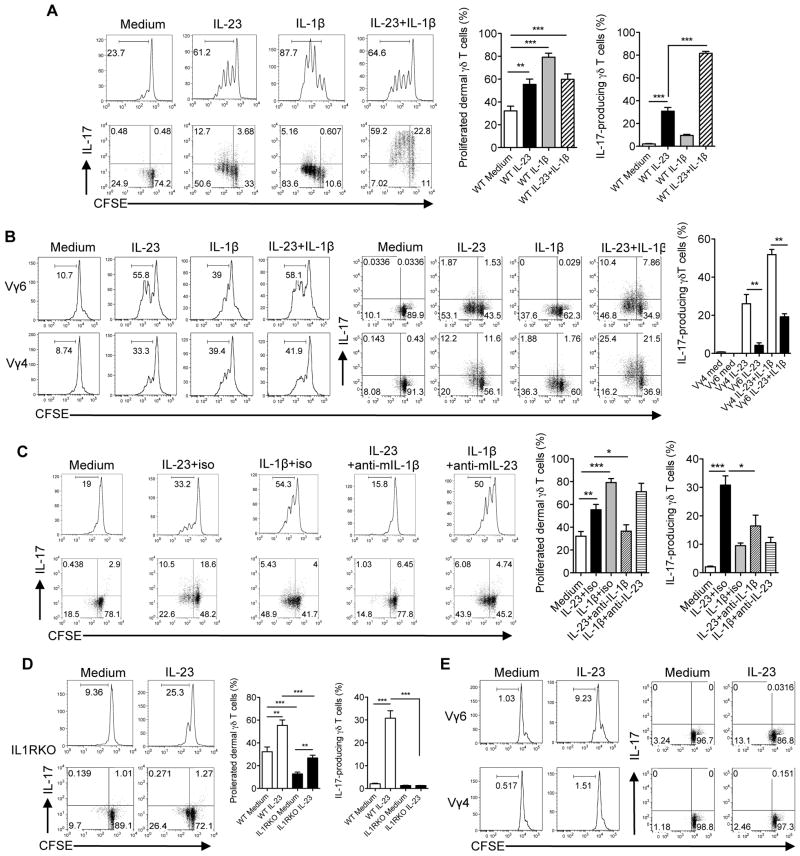
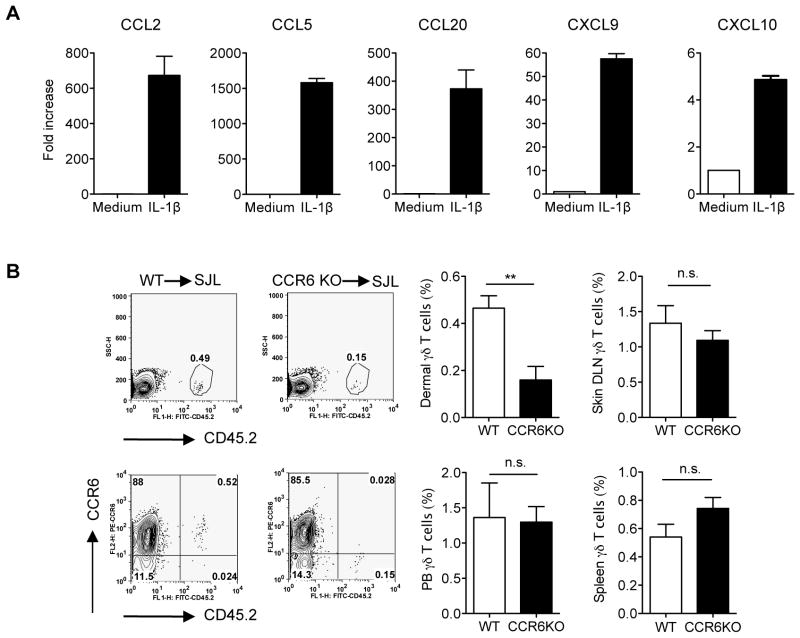
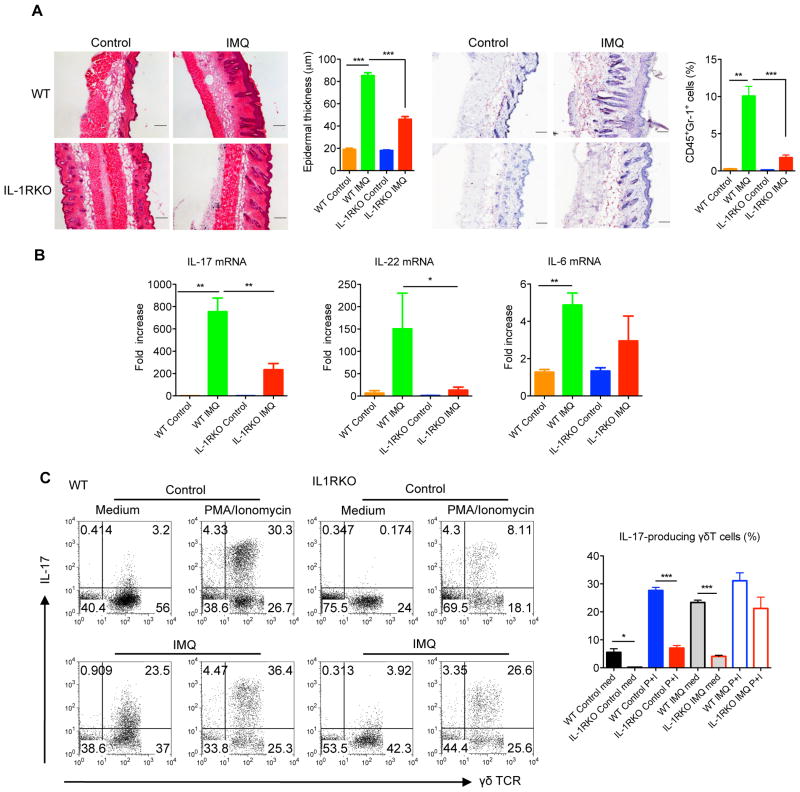
Dermal IL-17-producing γδT cells have a critical role in skin inflammation. However, their development and peripheral regulation have not been fully elucidated. Here we demonstrate that dermal γδT cells develop from the embryonic thymus and undergo homeostatic proliferation after birth with diversified TCR repertoire. Vγ6T cells are bona fide resident, but precursors of dermal Vγ4T cells may require extrathymic environment for imprinting skin-homing properties. Thymic Vγ6T cells are more competitive than Vγ4 for dermal γδT cell reconstitution and TCRδ(-/-) mice reconstituted with Vγ6 develop psoriasis-like inflammation after IMQ-application. Although both IL-23 and IL-1β promote Vγ4 and Vγ6 proliferation, Vγ4 are the main source of IL-17 production that requires IL-1 signalling. Mice with deficiency of IL-1RI signalling have significantly decreased skin inflammation. These studies reveal a differential developmental requirement and peripheral regulation for dermal Vγ6 and Vγ4 γδT cells, implying a new mechanism that may be involved in skin inflammation.
Figures
References
-
- Di Meglio P, Perera GK, Nestle FO. The multitasking organ: recent insights into skin immune function. Immunity. 2011;35:857–869. - PubMed
-
- Payer E, Elbe A, Stingl G. Epidermal T lymphocytes--ontogeny, features and function. Springer seminars in immunopathology. 1992;13:315–331. - PubMed
-
- Sharp LL, Jameson JM, Cauvi G, Havran WL. Dendritic epidermal T cells regulate skin homeostasis through local production of insulin-like growth factor 1. Nat Immunol. 2005;6:73–79. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
