

Activation of the aryl hydrocarbon receptor dampens the severity of inflammatory skin conditions
- PMID: 24909886
- PMCID: PMC4067745
- DOI: 10.1016/j.immuni.2014.04.019
Activation of the aryl hydrocarbon receptor dampens the severity of inflammatory skin conditions
Abstract
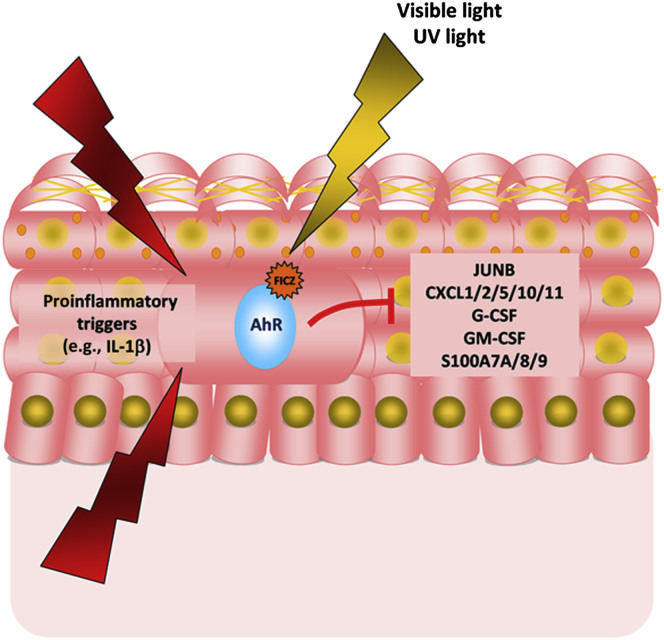
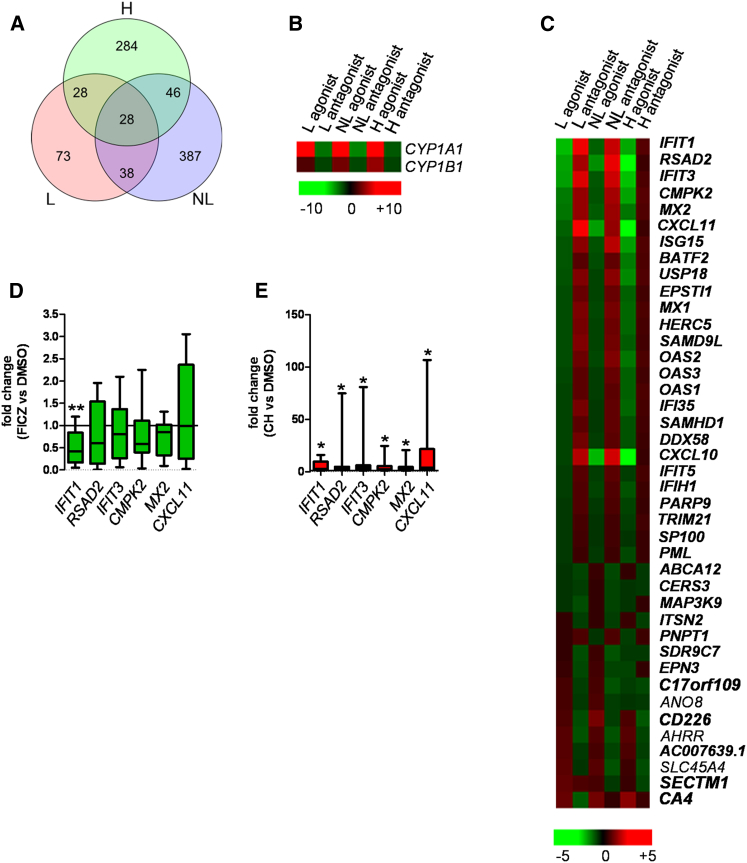
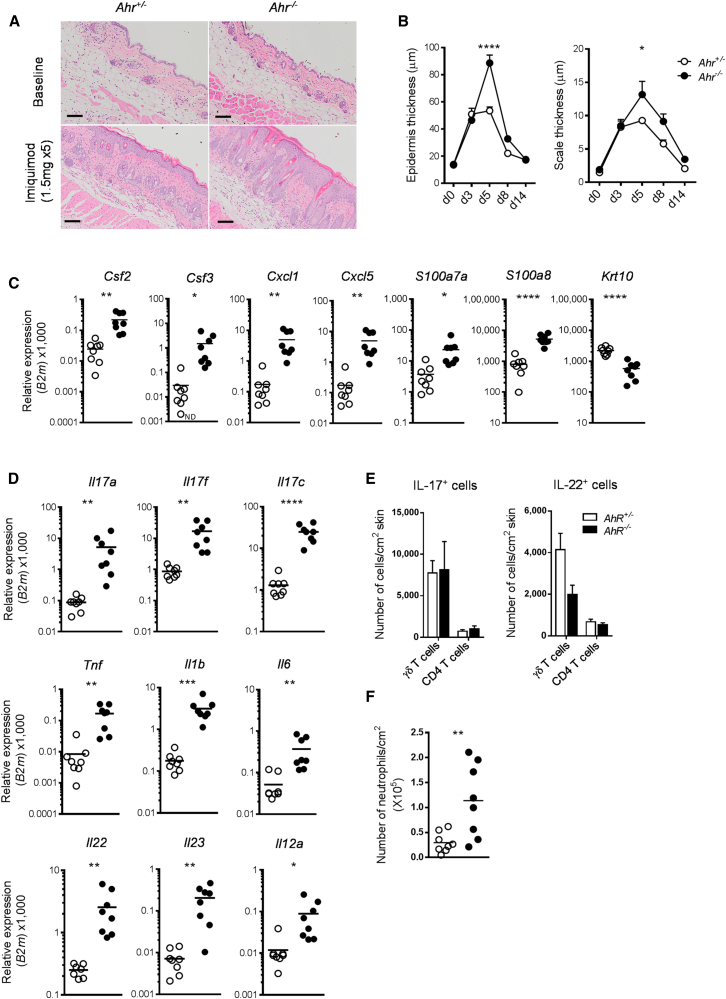
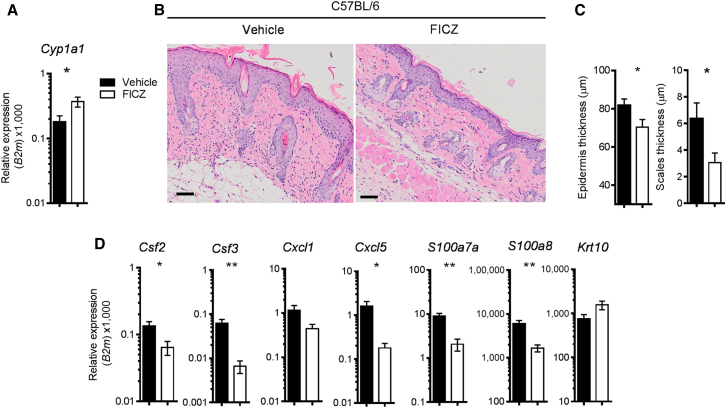
Environmental stimuli are known to contribute to psoriasis pathogenesis and that of other autoimmune diseases, but the mechanisms are largely unknown. Here we show that the aryl hydrocarbon receptor (AhR), a transcription factor that senses environmental stimuli, modulates pathology in psoriasis. AhR-activating ligands reduced inflammation in the lesional skin of psoriasis patients, whereas AhR antagonists increased inflammation. Similarly, AhR signaling via the endogenous ligand FICZ reduced the inflammatory response in the imiquimod-induced model of skin inflammation and AhR-deficient mice exhibited a substantial exacerbation of the disease, compared to AhR-sufficient controls. Nonhematopoietic cells, in particular keratinocytes, were responsible for this hyperinflammatory response, which involved upregulation of AP-1 family members of transcription factors. Thus, our data suggest a critical role for AhR in the regulation of inflammatory responses and open the possibility for novel therapeutic strategies in chronic inflammatory disorders.
Copyright © 2014 The Authors. Published by Elsevier Inc. All rights reserved.
Figures
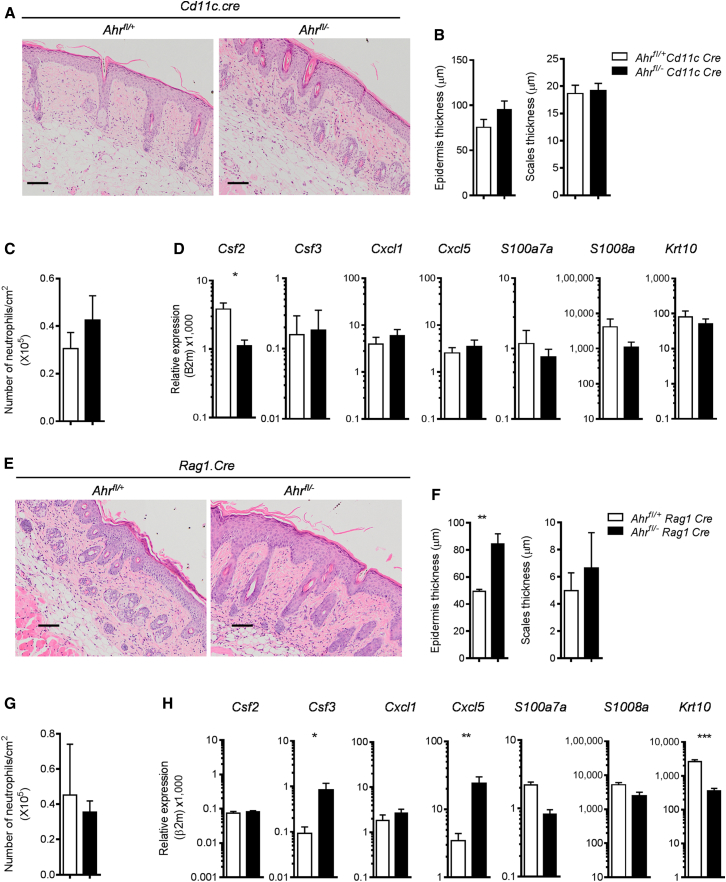
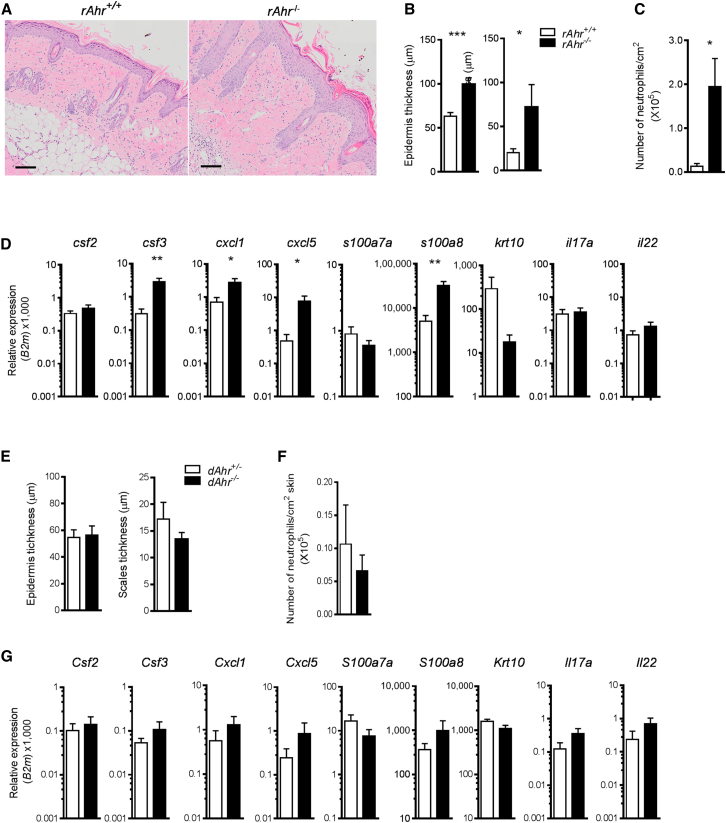
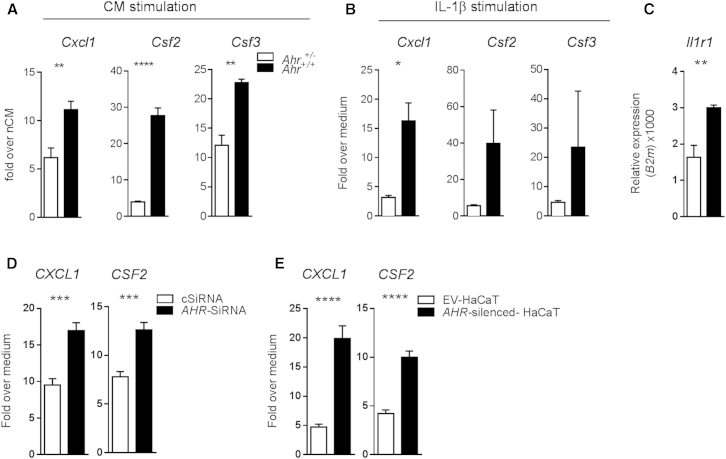
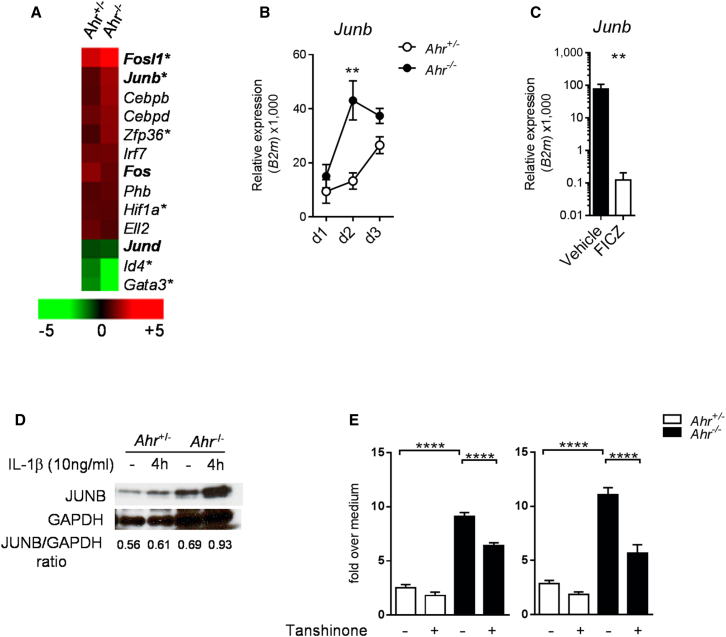
Comment in
-
AHR: making the keratinocytes thick skinned.Immunity. 2014 Jun 19;40(6):863-4. doi: 10.1016/j.immuni.2014.06.001. Immunity. 2014. PMID: 24950209
References
-
- Austin L.M., Ozawa M., Kikuchi T., Walters I.B., Krueger J.G. The majority of epidermal T cells in Psoriasis vulgaris lesions can produce type 1 cytokines, interferon-gamma, interleukin-2, and tumor necrosis factor-alpha, defining TC1 (cytotoxic T lymphocyte) and TH1 effector populations: a type 1 differentiation bias is also measured in circulating blood T cells in psoriatic patients. J. Invest. Dermatol. 1999;113:752–759. - PubMed
-
- Bata-Csorgo Z., Szell M. The psoriatic keratinocytes. Expert Rev. Dermatol. 2012;7:473–481.
-
- Bock K.W., Köhle C. Ah receptor: dioxin-mediated toxic responses as hints to deregulated physiologic functions. Biochem. Pharmacol. 2006;72:393–404. - PubMed
-
- Bowcock A.M., Shannon W., Du F., Duncan J., Cao K., Aftergut K., Catier J., Fernandez-Vina M.A., Menter A. Insights into psoriasis and other inflammatory diseases from large-scale gene expression studies. Hum. Mol. Genet. 2001;10:1793–1805. - PubMed
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
