

Direct observation of R-loop formation by single RNA-guided Cas9 and Cascade effector complexes
- PMID: 24912165
- PMCID: PMC4103346
- DOI: 10.1073/pnas.1402597111
Direct observation of R-loop formation by single RNA-guided Cas9 and Cascade effector complexes
Abstract
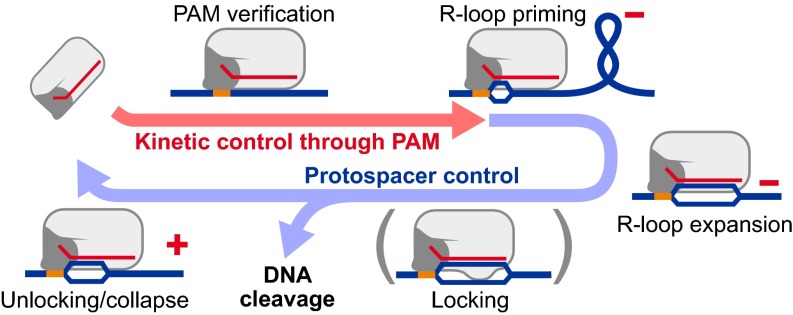
Clustered, regularly interspaced, short palindromic repeats (CRISPR)/CRISPR-associated (Cas) systems protect bacteria and archaea from infection by viruses and plasmids. Central to this defense is a ribonucleoprotein complex that produces RNA-guided cleavage of foreign nucleic acids. In DNA-targeting CRISPR-Cas systems, the RNA component of the complex encodes target recognition by forming a site-specific hybrid (R-loop) with its complement (protospacer) on an invading DNA while displacing the noncomplementary strand. Subsequently, the R-loop structure triggers DNA degradation. Although these reactions have been reconstituted, the exact mechanism of R-loop formation has not been fully resolved. Here, we use single-molecule DNA supercoiling to directly observe and quantify the dynamics of torque-dependent R-loop formation and dissociation for both Cascade- and Cas9-based CRISPR-Cas systems. We find that the protospacer adjacent motif (PAM) affects primarily the R-loop association rates, whereas protospacer elements distal to the PAM affect primarily R-loop stability. Furthermore, Cascade has higher torque stability than Cas9 by using a conformational locking step. Our data provide direct evidence for directional R-loop formation, starting from PAM recognition and expanding toward the distal protospacer end. Moreover, we introduce DNA supercoiling as a quantitative tool to explore the sequence requirements and promiscuities of orthogonal CRISPR-Cas systems in rapidly emerging gene-targeting applications.
Keywords: crRNA; genome engineering; magnetic tweezers.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
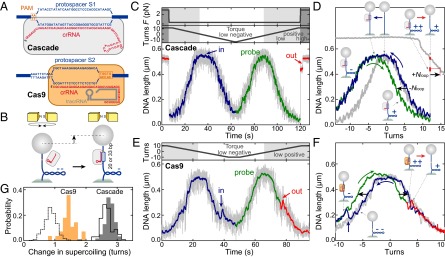
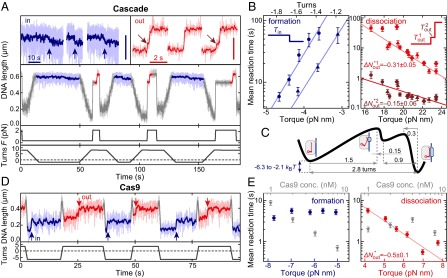
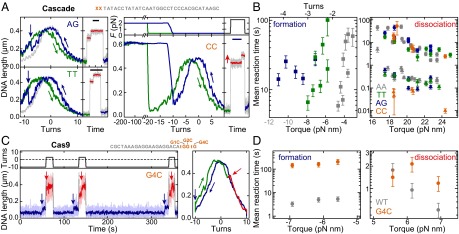
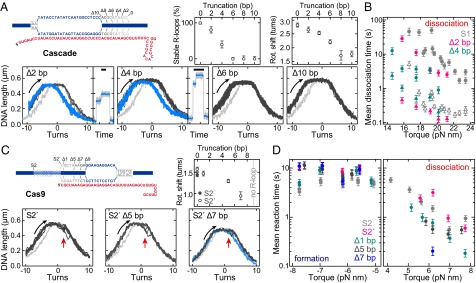
Comment in
-
Fulfilling the dream of a perfect genome editing tool.Proc Natl Acad Sci U S A. 2014 Jul 15;111(28):10029-30. doi: 10.1073/pnas.1408985111. Epub 2014 Jul 2. Proc Natl Acad Sci U S A. 2014. PMID: 24989506 Free PMC article. No abstract available.
References
-
- Garneau JE, et al. The CRISPR/Cas bacterial immune system cleaves bacteriophage and plasmid DNA. Nature. 2010;468(7320):67–71. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
