

An alternative, arginase-independent pathway for arginine metabolism in Kluyveromyces lactis involves guanidinobutyrase as a key enzyme
- PMID: 24912400
- PMCID: PMC4149782
- DOI: 10.1111/mmi.12666
An alternative, arginase-independent pathway for arginine metabolism in Kluyveromyces lactis involves guanidinobutyrase as a key enzyme
Abstract
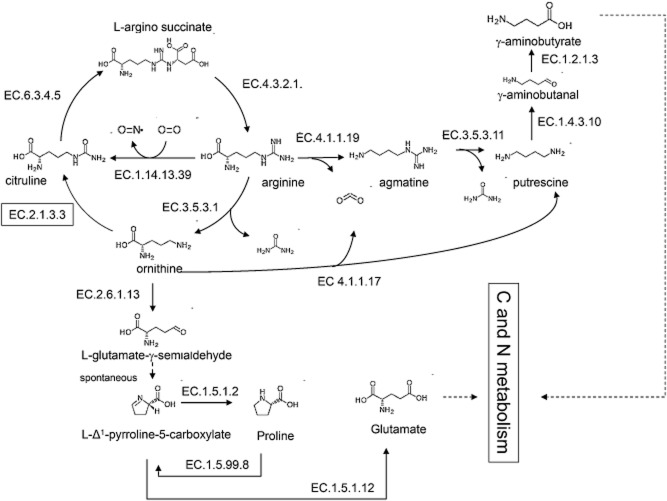
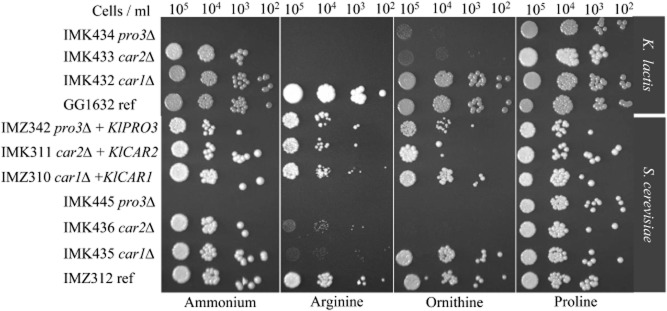
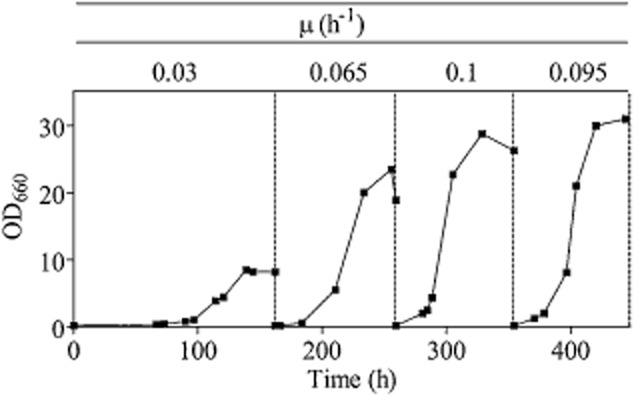
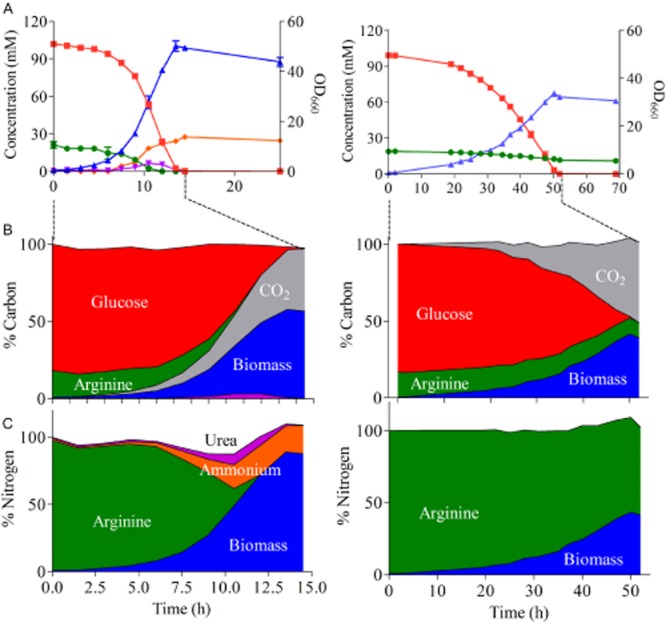
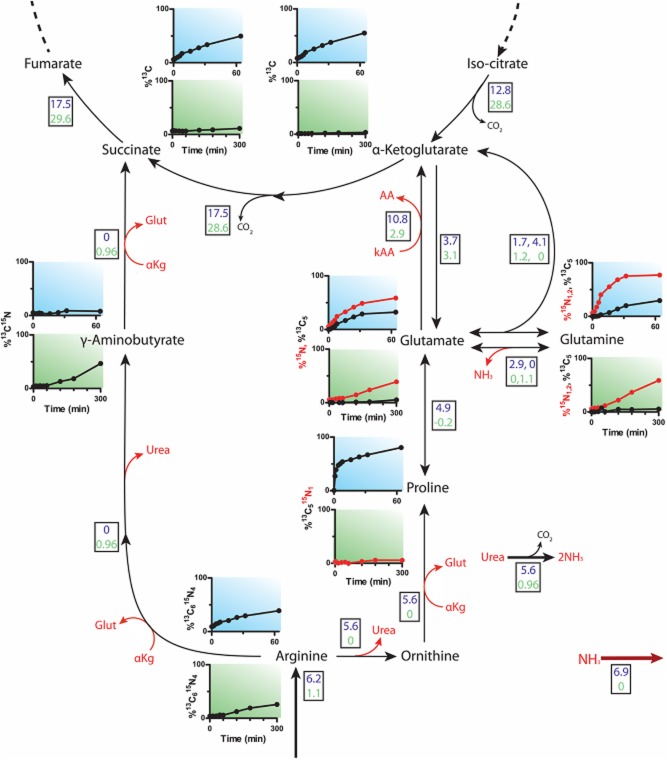
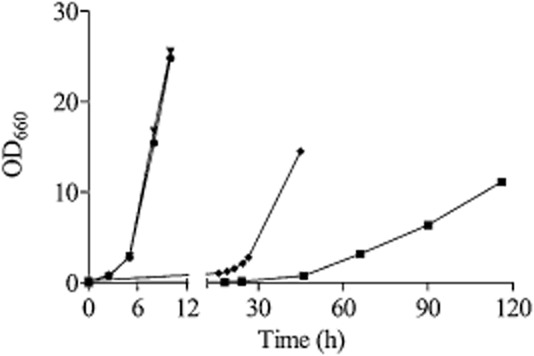
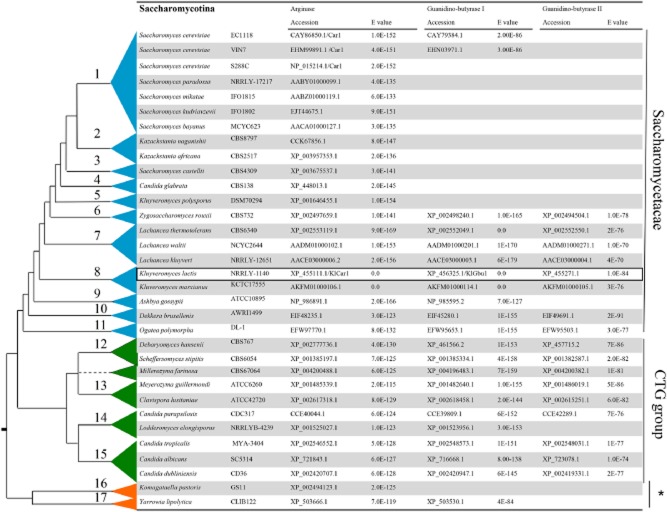
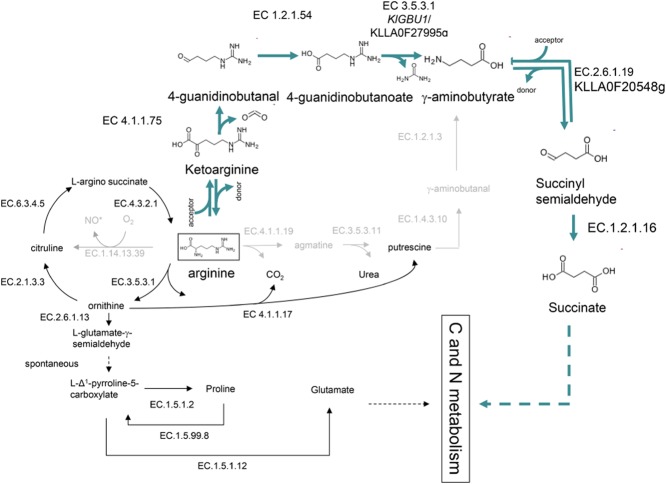
Most available knowledge on fungal arginine metabolism is derived from studies on Saccharomyces cerevisiae, in which arginine catabolism is initiated by releasing urea via the arginase reaction. Orthologues of the S. cerevisiae genes encoding the first three enzymes in the arginase pathway were cloned from Kluyveromyces lactis and shown to functionally complement the corresponding deletion in S. cerevisiae. Surprisingly, deletion of the single K. lactis arginase gene KlCAR1 did not completely abolish growth on arginine as nitrogen source. Growth rate of the deletion mutant strongly increased during serial transfer in shake-flask cultures. A combination of RNAseq-based transcriptome analysis and (13)C-(15)N-based flux analysis was used to elucidate the arginase-independent pathway. Isotopic (13)C(15)N-enrichment in γ-aminobutyrate revealed succinate as the entry point in the TCA cycle of the alternative pathway. Transcript analysis combined with enzyme activity measurements indicated increased expression in the Klcar1Δ mutant of a guanidinobutyrase (EC.3.5.3.7), a key enzyme in a new pathway for arginine degradation. Expression of the K. lactis KLLA0F27995g (renamed KlGBU1) encoding guanidinobutyrase enabled S. cerevisiae to use guanidinobutyrate as sole nitrogen source and its deletion in K. lactis almost completely abolish growth on this nitrogen source. Phylogenetic analysis suggests that this enzyme activity is widespread in fungi.
© 2014 The Authors. Molecular Microbiology published by John Wiley & Sons Ltd.
Figures
Similar articles
-
Three Related Enzymes in Candida albicans Achieve Arginine- and Agmatine-Dependent Metabolism That Is Essential for Growth and Fungal Virulence.mBio. 2020 Aug 11;11(4):e01845-20. doi: 10.1128/mBio.01845-20. mBio. 2020. PMID: 32788384 Free PMC article.
-
Cloning and sequencing of Schizosaccharomyces pombe car1 gene encoding arginase. Expression of the arginine anabolic and catabolic genes in response to arginine and related metabolites.Yeast. 1994 Jul;10(7):923-33. doi: 10.1002/yea.320100707. Yeast. 1994. PMID: 7985419
-
Differences in regulation of yeast gluconeogenesis revealed by Cat8p-independent activation of PCK1 and FBP1 genes in Kluyveromyces lactis.Mol Gen Genet. 2000 Sep;264(1-2):193-203. doi: 10.1007/s004380000314. Mol Gen Genet. 2000. PMID: 11016849
-
The deletion of the succinate dehydrogenase gene KlSDH1 in Kluyveromyces lactis does not lead to respiratory deficiency.Eukaryot Cell. 2004 Jun;3(3):589-97. doi: 10.1128/EC.3.3.589-597.2004. Eukaryot Cell. 2004. PMID: 15189981 Free PMC article.
-
Human arginase 1, a Jack of all trades?3 Biotech. 2022 Oct;12(10):264. doi: 10.1007/s13205-022-03326-9. Epub 2022 Sep 7. 3 Biotech. 2022. PMID: 36082360 Free PMC article. Review.
Cited by
-
Vitreous metabolomic signatures of pathological myopia with complications.Eye (Lond). 2023 Oct;37(14):2987-2993. doi: 10.1038/s41433-023-02457-4. Epub 2023 Feb 25. Eye (Lond). 2023. PMID: 36841867 Free PMC article.
-
Respiratory reoxidation of NADH is a key contributor to high oxygen requirements of oxygen-limited cultures of Ogataea parapolymorpha.FEMS Yeast Res. 2022 Feb 22;22(1):foac007. doi: 10.1093/femsyr/foac007. FEMS Yeast Res. 2022. PMID: 35137036 Free PMC article.
-
Mitochondrial Metabolomics of Sym1-Depleted Yeast Cells Revealed Them to Be Lysine Auxotroph.Cells. 2023 Feb 22;12(5):692. doi: 10.3390/cells12050692. Cells. 2023. PMID: 36899826 Free PMC article.
-
Examining gut microbiota and metabolites to clarify mechanisms of Dimocarpus longan Lour leaf components against type 2 diabetes.World J Diabetes. 2025 Jul 15;16(7):104512. doi: 10.4239/wjd.v16.i7.104512. World J Diabetes. 2025. PMID: 40697601 Free PMC article.
-
Three Related Enzymes in Candida albicans Achieve Arginine- and Agmatine-Dependent Metabolism That Is Essential for Growth and Fungal Virulence.mBio. 2020 Aug 11;11(4):e01845-20. doi: 10.1128/mBio.01845-20. mBio. 2020. PMID: 32788384 Free PMC article.
References
-
- Abdelal AT. Arginine catabolism by microorganisms. Annu Rev Microbiol. 1979;33:139–168. - PubMed
-
- Archibald RM. Colorimetric determination of urea. J Biol Chem. 1945;157:507–518.
-
- Beltran G, Esteve-Zarzoso B, Rozes N, Mas A, Guillamon JM. Influence of the timing of nitrogen additions during synthetic grape must fermentations on fermentation kinetics and nitrogen consumption. J Agric Food Chem. 2005;53:996–1002. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
