

A novel proteasome inhibitor suppresses tumor growth via targeting both 19S proteasome deubiquitinases and 20S proteolytic peptidases
- PMID: 24912524
- PMCID: PMC4050382
- DOI: 10.1038/srep05240
A novel proteasome inhibitor suppresses tumor growth via targeting both 19S proteasome deubiquitinases and 20S proteolytic peptidases
Abstract
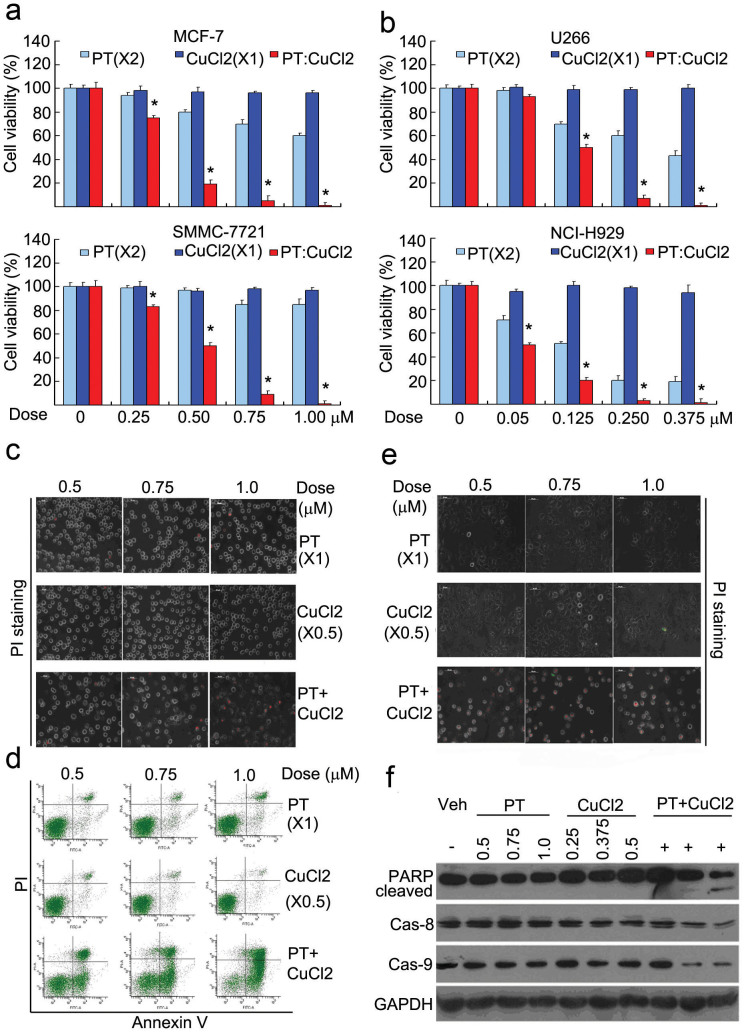
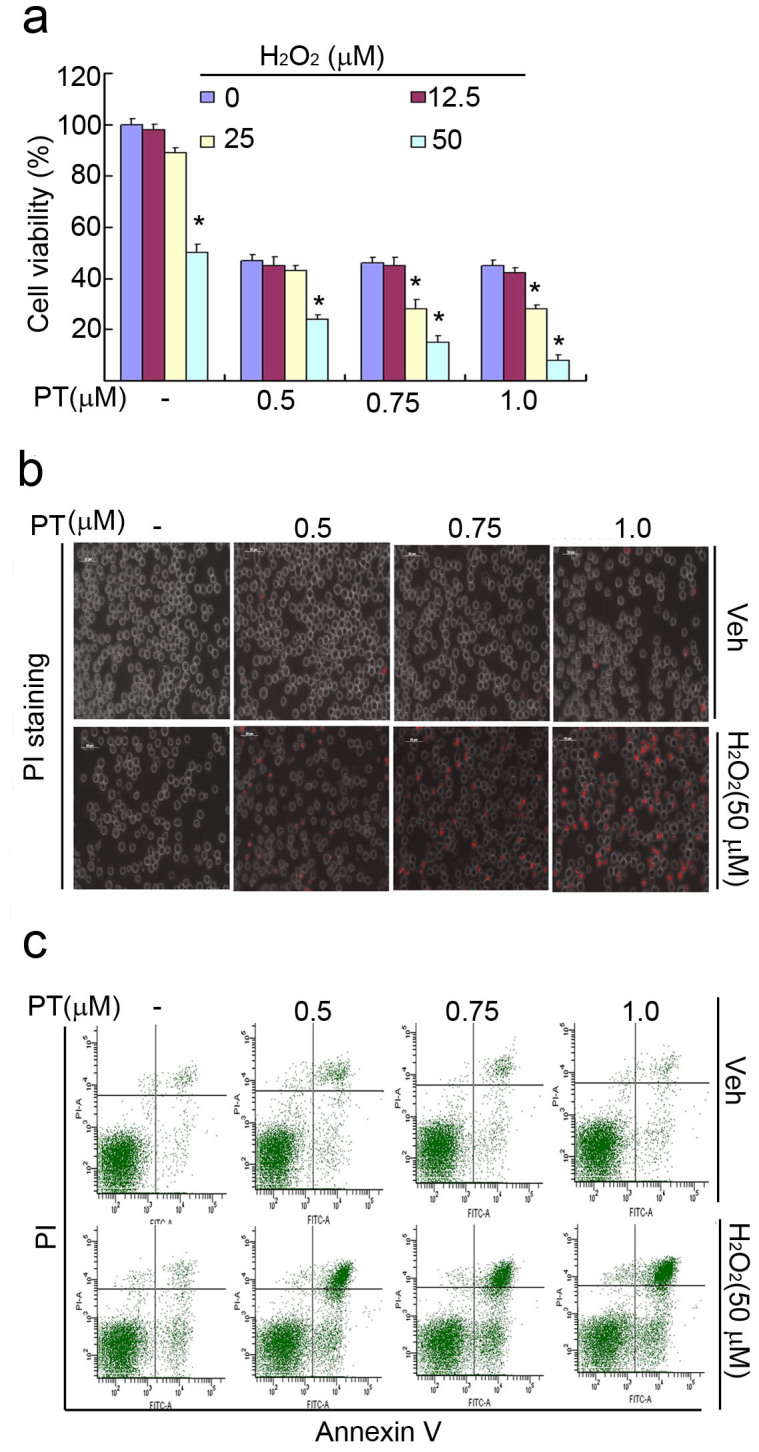
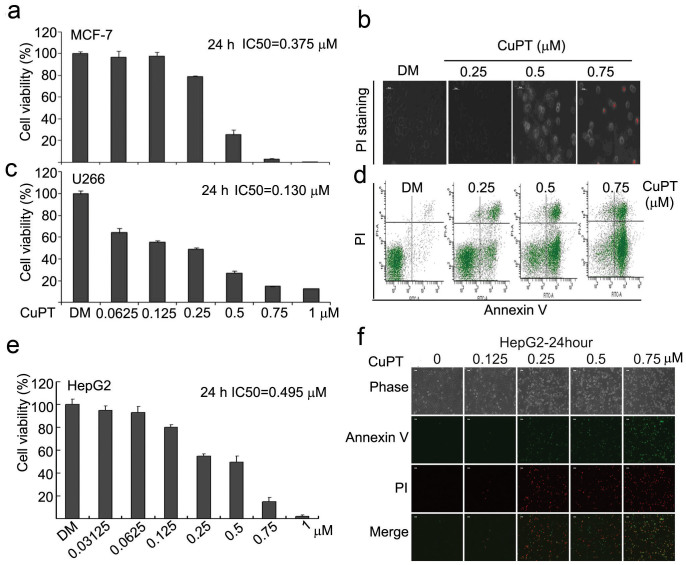
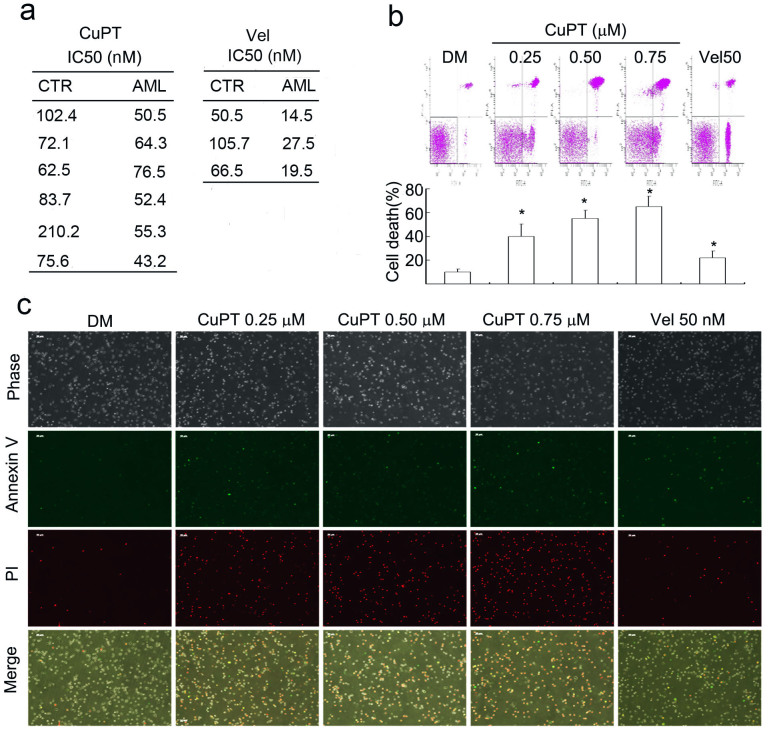
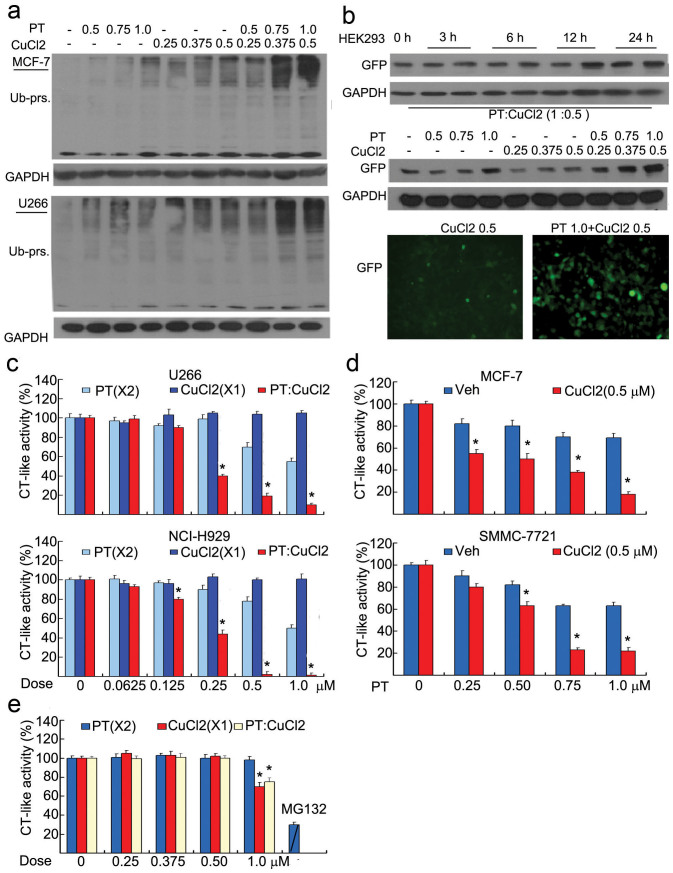
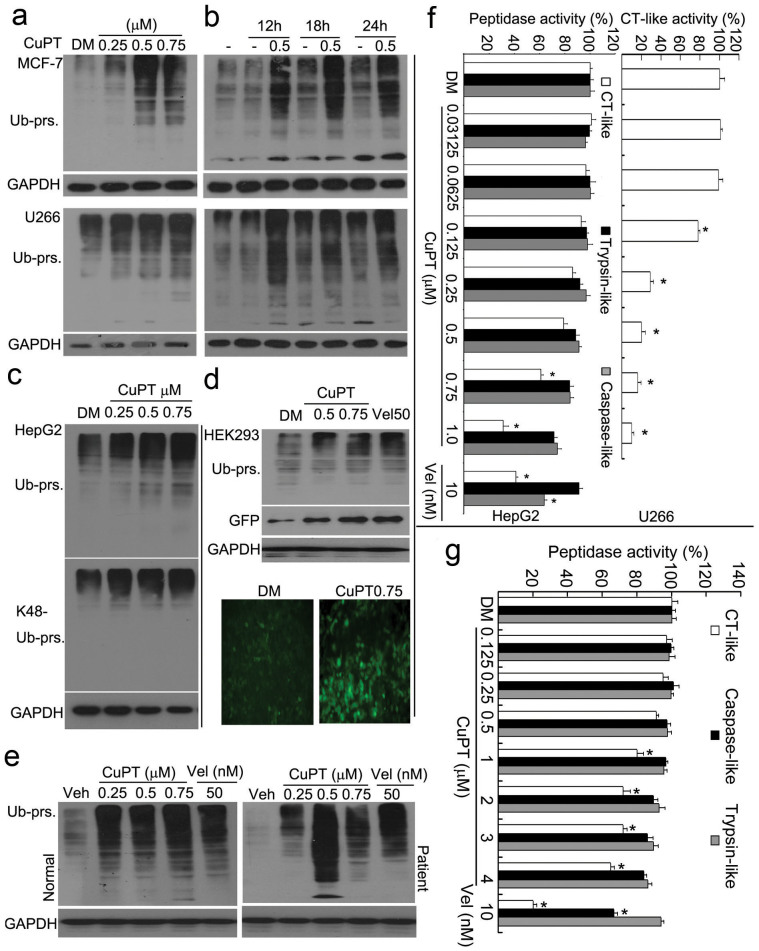
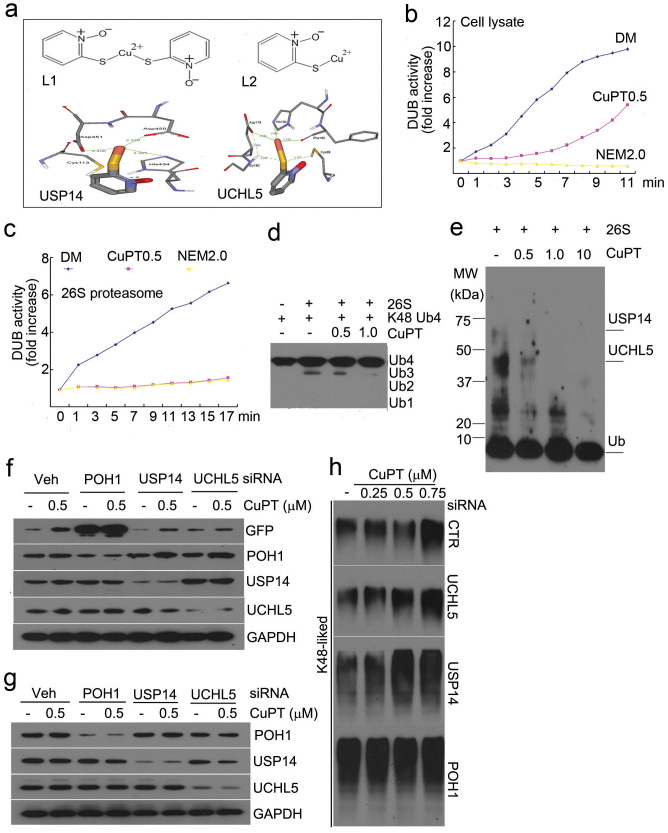
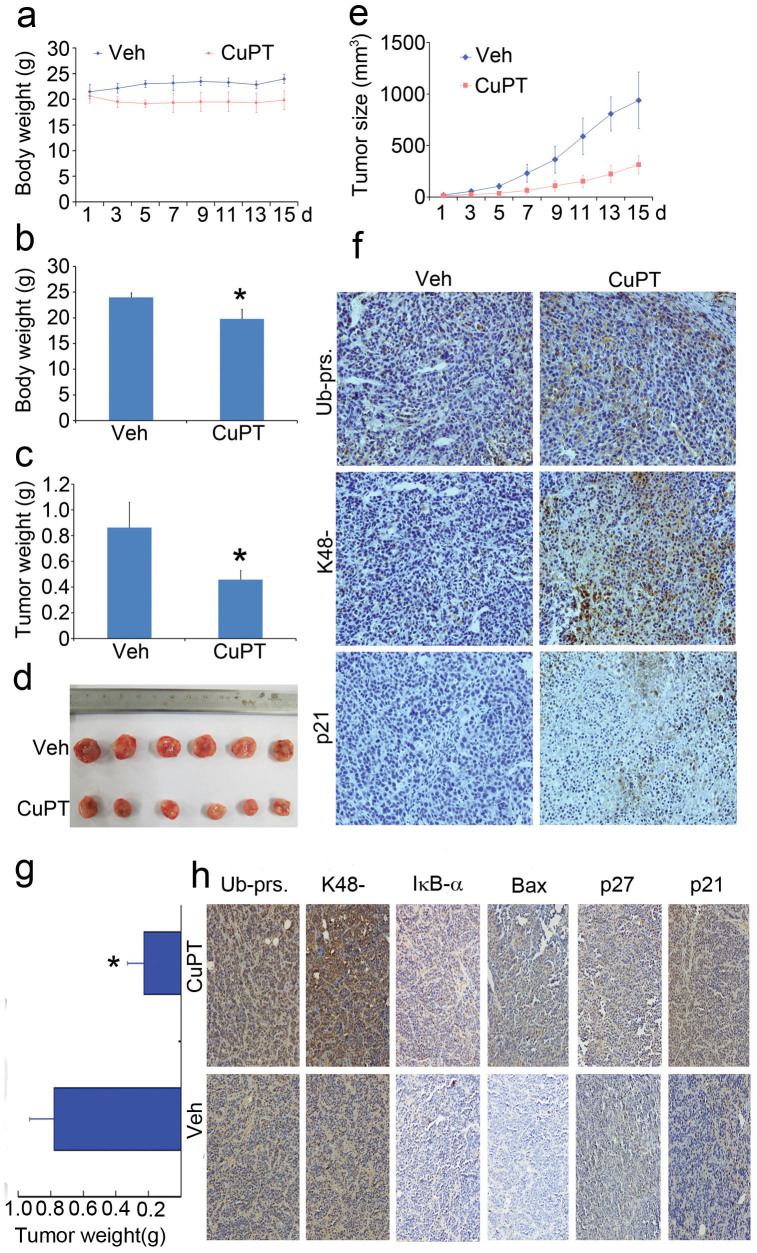
The successful development of bortezomib-based therapy for treatment of multiple myeloma has established proteasome inhibition as an effective therapeutic strategy, and both 20S proteasome peptidases and 19S deubiquitinases (DUBs) are becoming attractive targets of cancer therapy. It has been reported that metal complexes, such as copper complexes, inhibit tumor proteasome. However, the involved mechanism of action has not been fully characterized. Here we report that (i) copper pyrithione (CuPT), an alternative to tributyltin for antifouling paint biocides, inhibits the ubiquitin-proteasome system (UPS) via targeting both 19S proteasome-specific DUBs and 20S proteolytic peptidases with a mechanism distinct from that of the FDA-approved proteasome inhibitor bortezomib; (ii) CuPT potently inhibits proteasome-specific UCHL5 and USP14 activities; (iii) CuPT inhibits tumor growth in vivo and induces cytotoxicity in vitro and ex vivo. This study uncovers a novel class of dual inhibitors of DUBs and proteasome and suggests a potential clinical strategy for cancer therapy.
Figures
References
-
- Richardson P. G. et al. A phase 2 study of bortezomib in relapsed, refractory myeloma. N. Engl. J. Med. 348, 2609–2617 (2003). - PubMed
-
- Adams J. The development of proteasome inhibitors as anticancer drugs. Cancer Cell 5, 417–421 (2004). - PubMed
-
- Richardson P. G. et al. Bortezomib or high-dose dexamethasone for relapsed multiple myeloma. N. Engl. J. Med. 352, 2487–2498 (2005). - PubMed
-
- Galanski M., Arion V. B., Jakupec M. A. & Keppler B. K. Recent developments in the field of tumor-inhibiting metal complexes. Curr. Pharm. Des. 9, 2078–2089 (2003). - PubMed
-
- Galanski M., Jakupec M. A. & Keppler B. K. Update of the preclinical situation of anticancer platinum complexes: novel design strategies and innovative analytical approaches. Curr. Med. Chem. 12, 2075–2094 (2005). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
