

Age-dependent transcriptome and proteome following transection of neonatal spinal cord of Monodelphis domestica (South American grey short-tailed opossum)
- PMID: 24914927
- PMCID: PMC4051688
- DOI: 10.1371/journal.pone.0099080
Age-dependent transcriptome and proteome following transection of neonatal spinal cord of Monodelphis domestica (South American grey short-tailed opossum)
Abstract
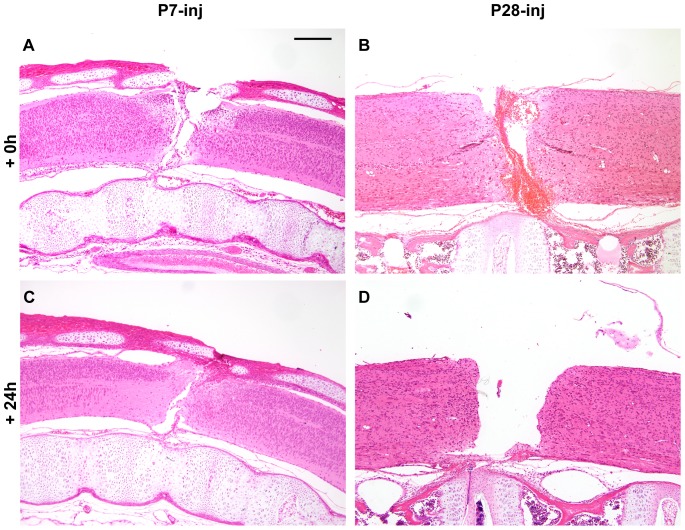
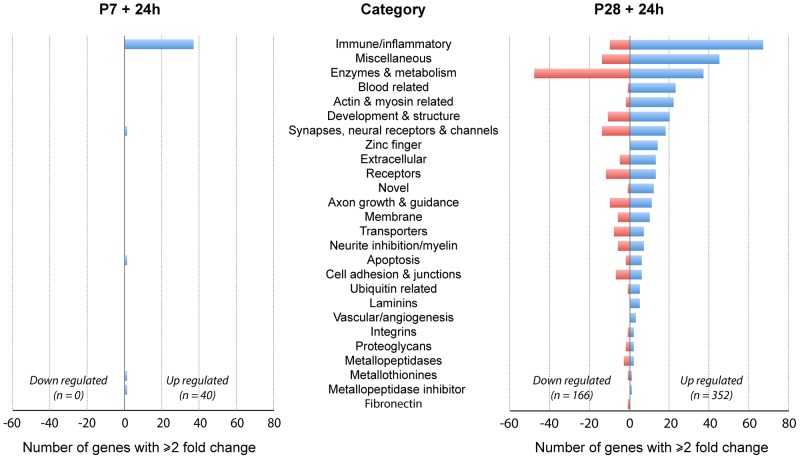
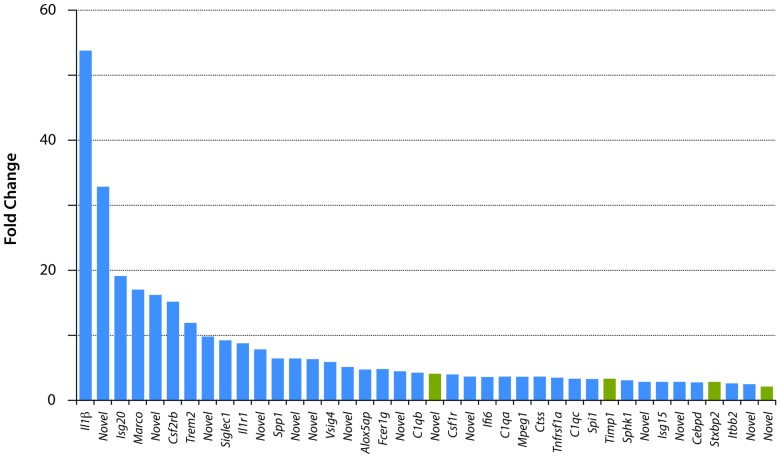
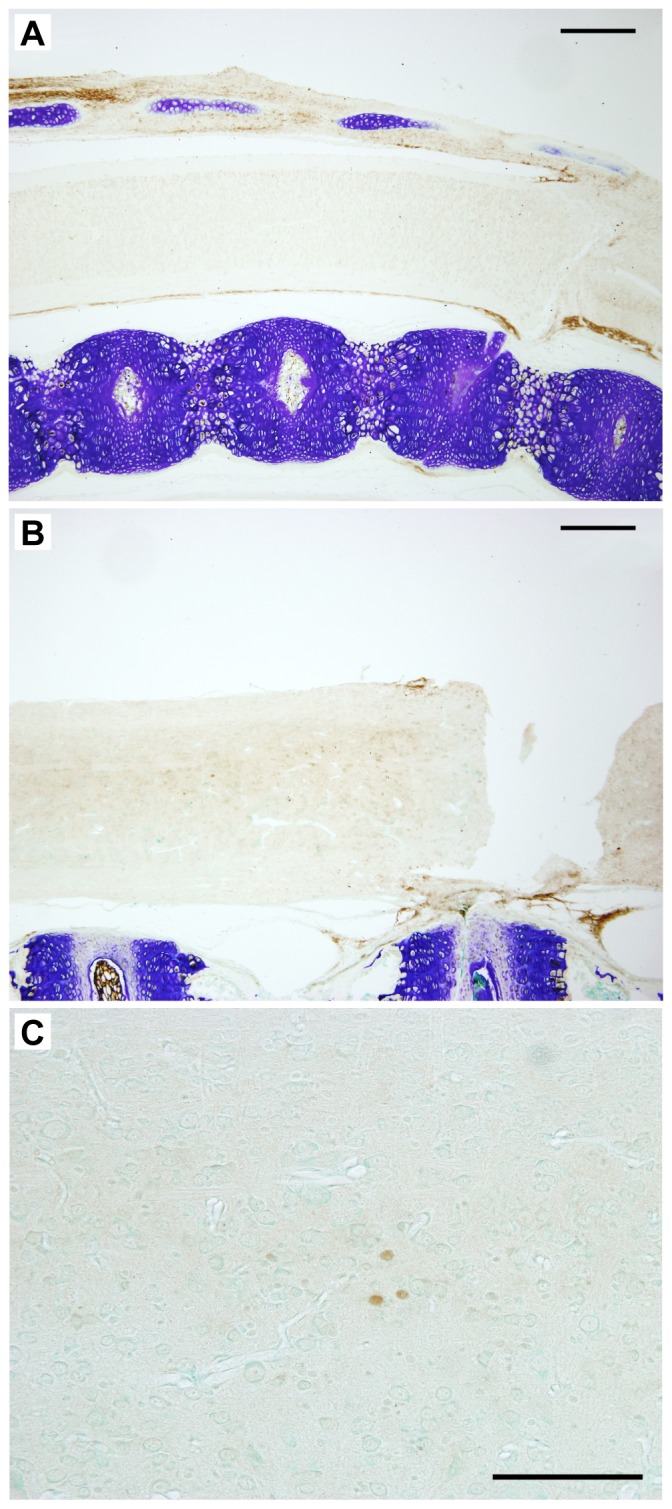
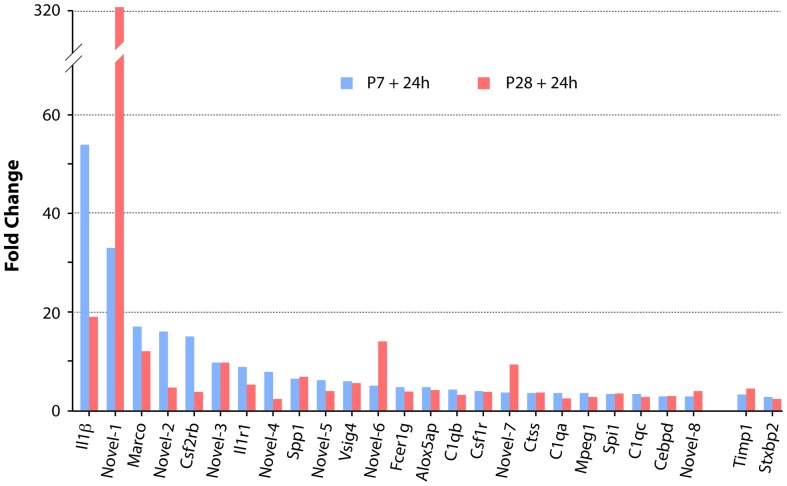
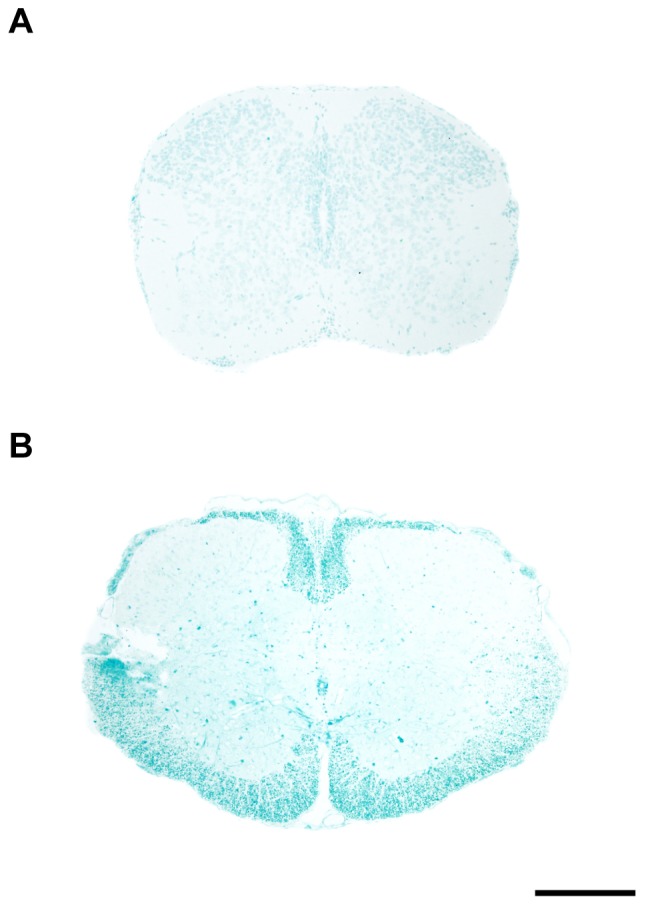
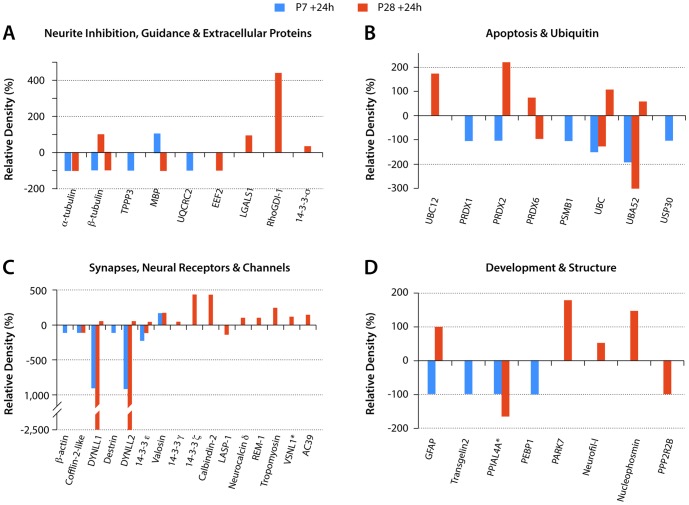
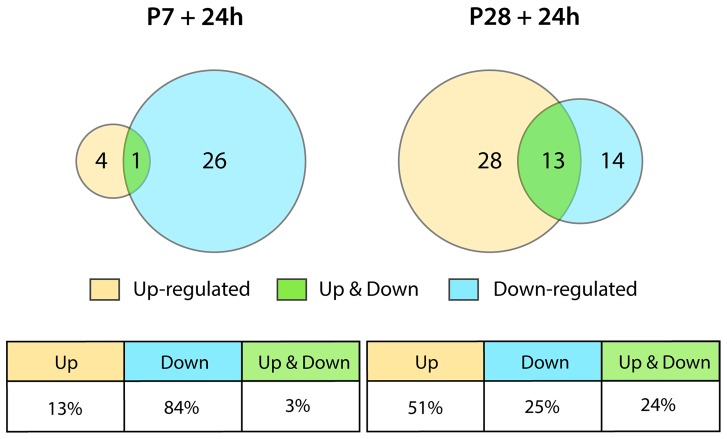
This study describes a combined transcriptome and proteome analysis of Monodelphis domestica response to spinal cord injury at two different postnatal ages. Previously we showed that complete transection at postnatal day 7 (P7) is followed by profuse axon growth across the lesion with near-normal locomotion and swimming when adult. In contrast, at P28 there is no axon growth across the lesion, the animals exhibit weight-bearing locomotion, but cannot use hind limbs when swimming. Here we examined changes in gene and protein expression in the segment of spinal cord rostral to the lesion at 24 h after transection at P7 and at P28. Following injury at P7 only forty genes changed (all increased expression); most were immune/inflammatory genes. Following injury at P28 many more genes changed their expression and the magnitude of change for some genes was strikingly greater. Again many were associated with the immune/inflammation response. In functional groups known to be inhibitory to regeneration in adult cords the expression changes were generally muted, in some cases opposite to that required to account for neurite inhibition. For example myelin basic protein expression was reduced following injury at P28 both at the gene and protein levels. Only four genes from families with extracellular matrix functions thought to influence neurite outgrowth in adult injured cords showed substantial changes in expression following injury at P28: Olfactomedin 4 (Olfm4, 480 fold compared to controls), matrix metallopeptidase (Mmp1, 104 fold), papilin (Papln, 152 fold) and integrin α4 (Itga4, 57 fold). These data provide a resource for investigation of a priori hypotheses in future studies of mechanisms of spinal cord regeneration in immature animals compared to lack of regeneration at more mature stages.
Conflict of interest statement
Figures
References
-
- Aguayo AJ, David S, Bray GM (1981) Influences of the glial environment on the elongation of axons after injury: transplantation studies in adult rodents. The Journal of experimental biology 95: 231–240. - PubMed
-
- David S, Aguayo AJ (1981) Axonal elongation into peripheral nervous system "bridges" after central nervous system injury in adult rats. Science 214: 931–933. - PubMed
-
- Tello F (1911) La influencia del neurotropismo en la regeneracion de los centros nerviosos. Trab Laborator Invest Biol University Madrid 9: 123–159.
-
- Buchli AD, Schwab ME (2005) Inhibition of Nogo: a key strategy to increase regeneration, plasticity and functional recovery of the lesioned central nervous system. Annals of medicine 37: 556–567. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
