

Structure of the phosphotransferase domain of the bifunctional aminoglycoside-resistance enzyme AAC(6')-Ie-APH(2'')-Ia
- PMID: 24914967
- PMCID: PMC4051501
- DOI: 10.1107/S1399004714005331
Structure of the phosphotransferase domain of the bifunctional aminoglycoside-resistance enzyme AAC(6')-Ie-APH(2'')-Ia
Abstract
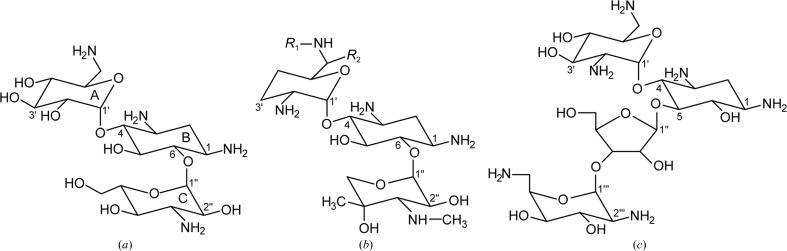
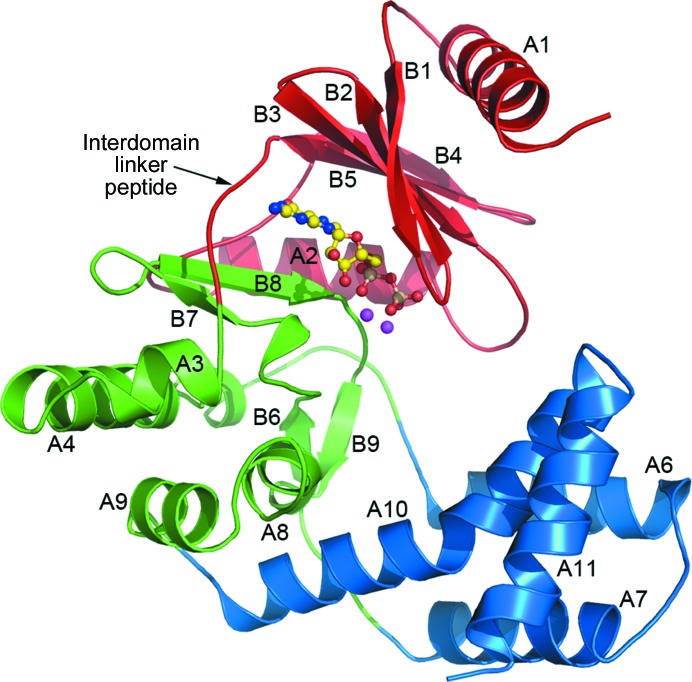
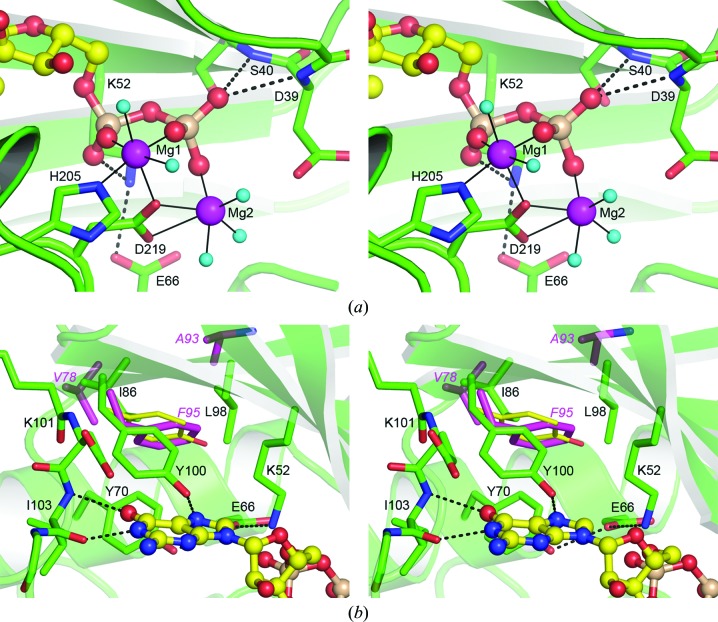
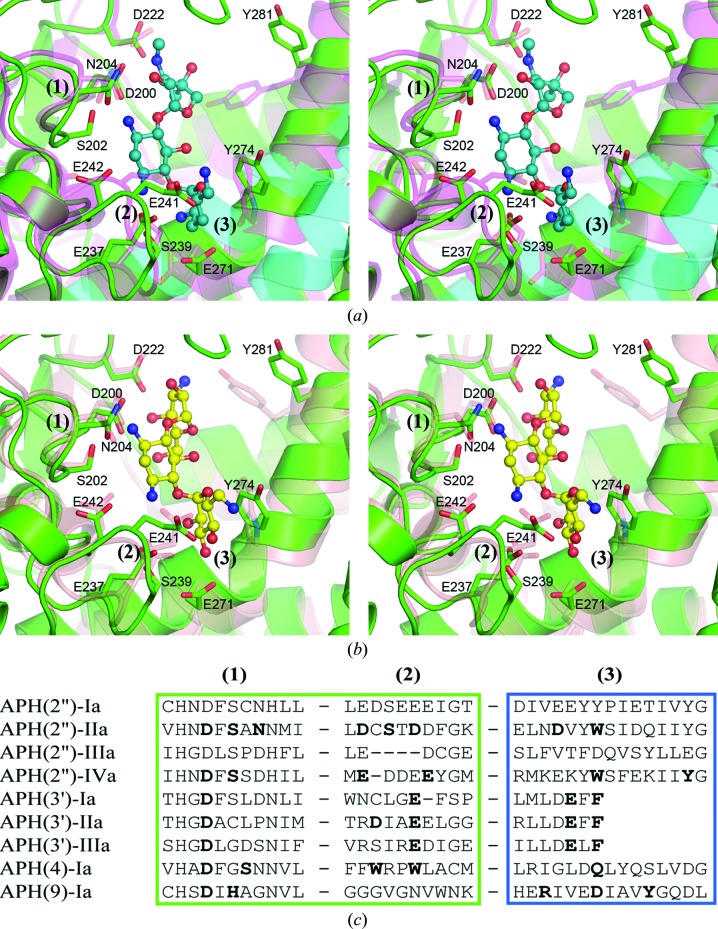
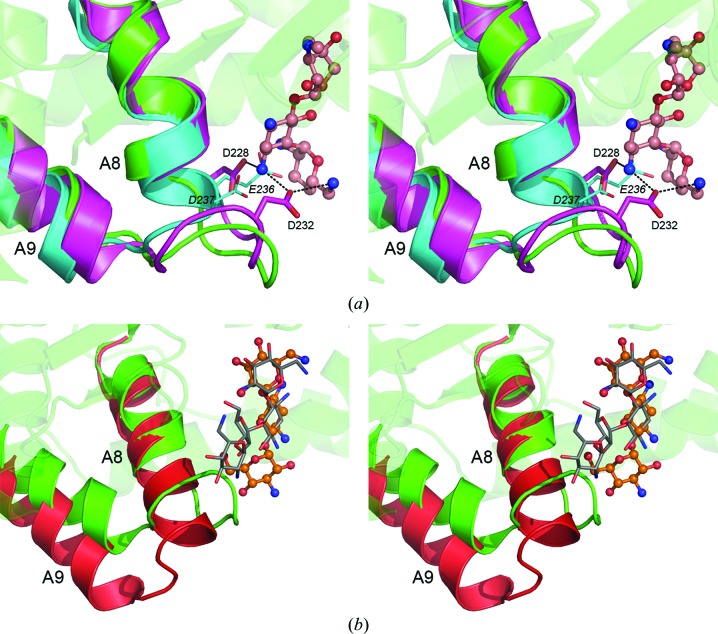
The bifunctional acetyltransferase(6')-Ie-phosphotransferase(2'')-Ia [AAC(6')-Ie-APH(2'')-Ia] is the most important aminoglycoside-resistance enzyme in Gram-positive bacteria, conferring resistance to almost all known aminoglycoside antibiotics in clinical use. Owing to its importance, this enzyme has been the focus of intensive research since its isolation in the mid-1980s but, despite much effort, structural details of AAC(6')-Ie-APH(2'')-Ia have remained elusive. The structure of the Mg2GDP complex of the APH(2'')-Ia domain of the bifunctional enzyme has now been determined at 2.3 Å resolution. The structure of APH(2'')-Ia is reminiscent of the structures of other aminoglycoside phosphotransferases, having a two-domain architecture with the nucleotide-binding site located at the junction of the two domains. Unlike the previously characterized APH(2'')-IIa and APH(2'')-IVa enzymes, which are capable of utilizing both ATP and GTP as the phosphate donors, APH(2'')-Ia uses GTP exclusively in the phosphorylation of the aminoglycoside antibiotics, and in this regard closely resembles the GTP-dependent APH(2'')-IIIa enzyme. In APH(2'')-Ia this GTP selectivity is governed by the presence of a `gatekeeper' residue, Tyr100, the side chain of which projects into the active site and effectively blocks access to the adenine-binding template. Mutation of this tyrosine residue to a less bulky phenylalanine provides better access for ATP to the NTP-binding template and converts APH(2'')-Ia into a dual-specificity enzyme.
Keywords: AAC(6′)-Ie-APH(2′′)-Ia; aminoglycoside-resistance enzymes.
Figures
References
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
