

Laminin α2-mediated focal adhesion kinase activation triggers Alport glomerular pathogenesis
- PMID: 24915008
- PMCID: PMC4051676
- DOI: 10.1371/journal.pone.0099083
Laminin α2-mediated focal adhesion kinase activation triggers Alport glomerular pathogenesis
Abstract
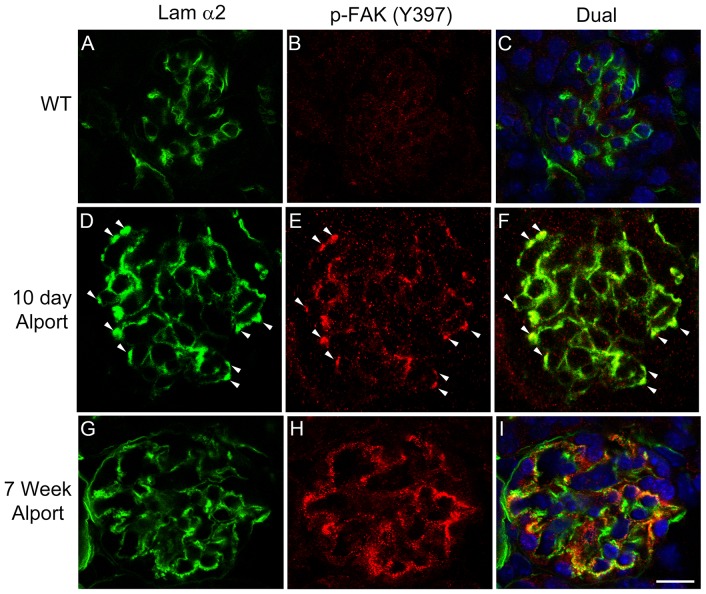
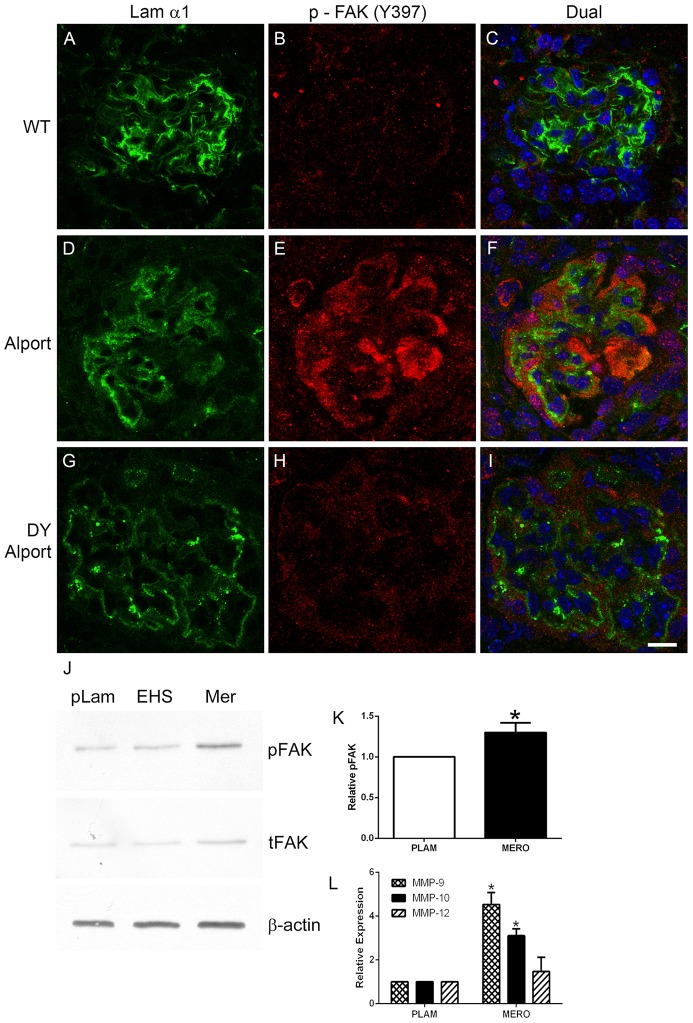
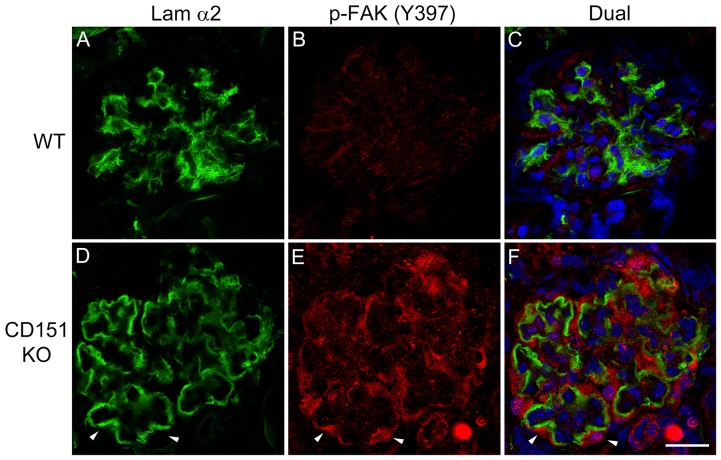
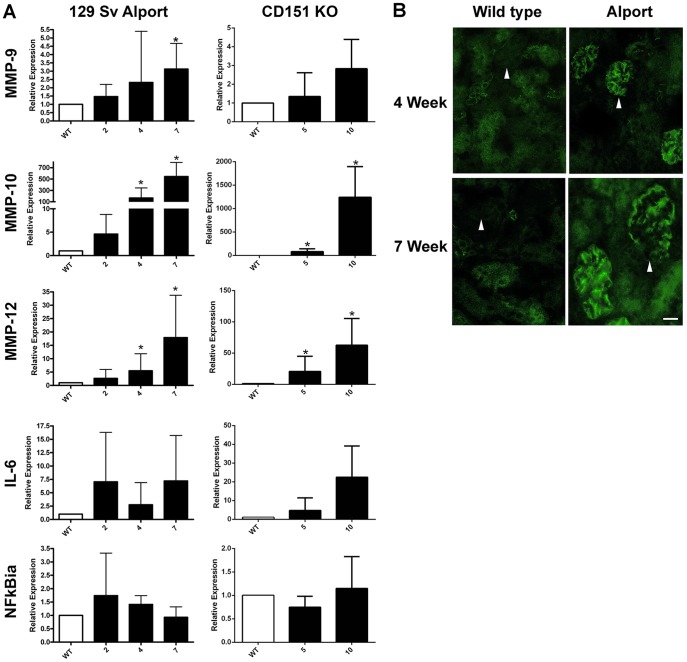
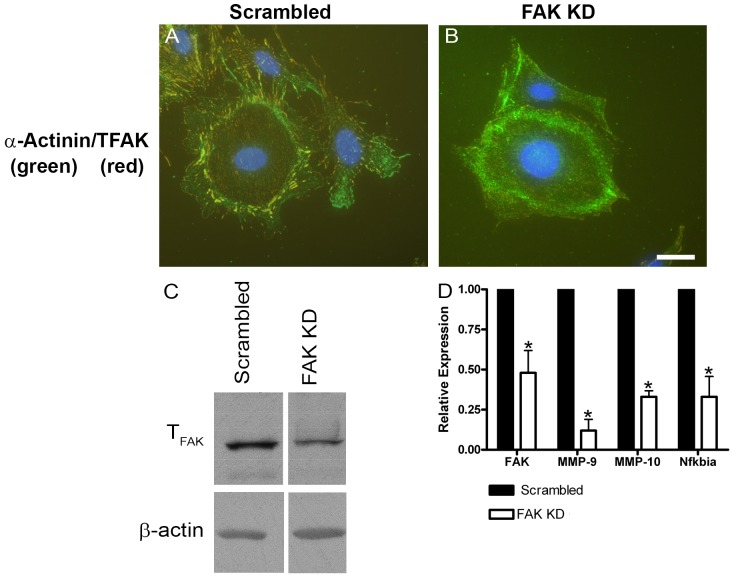
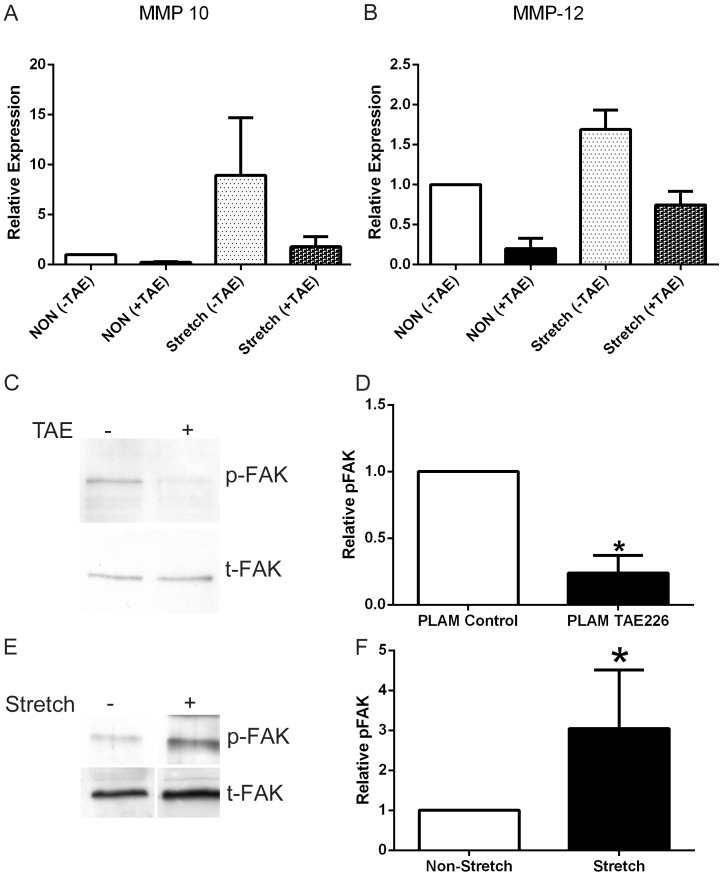
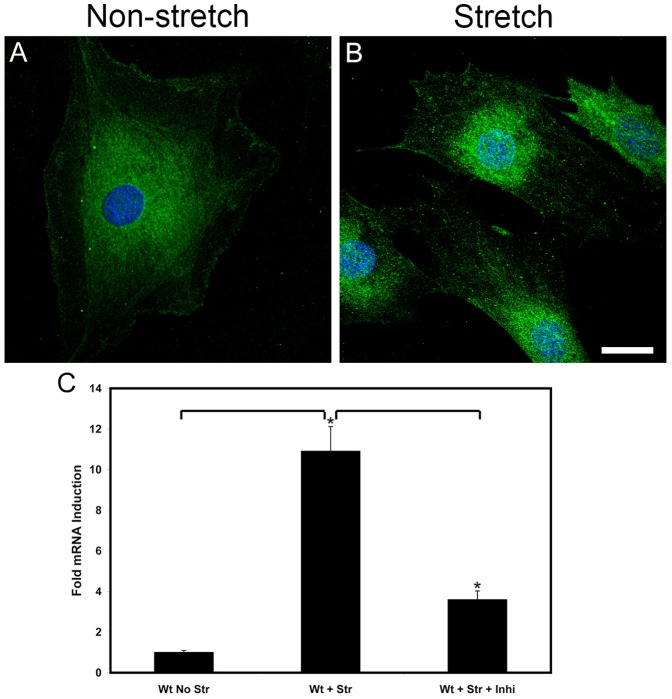
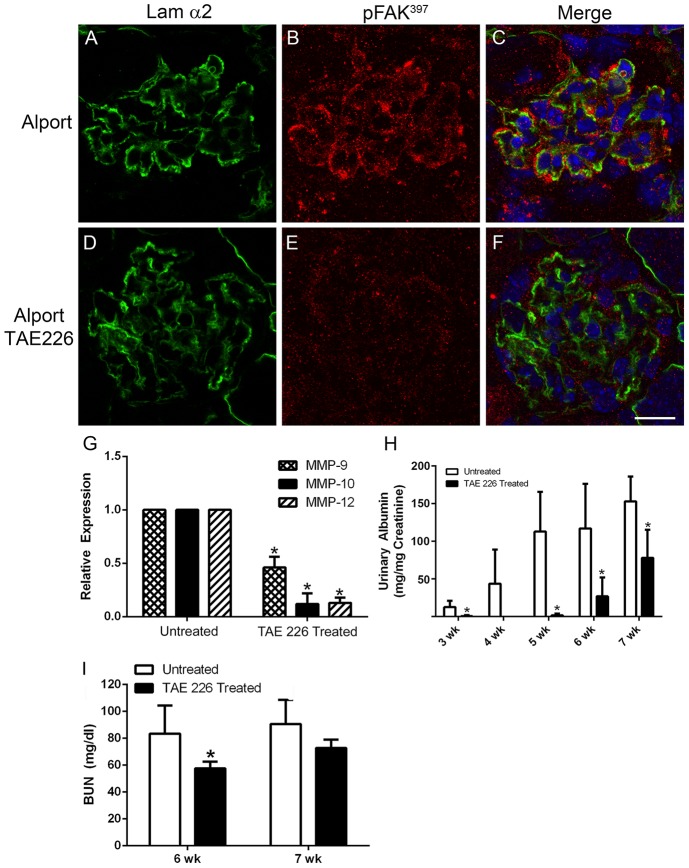
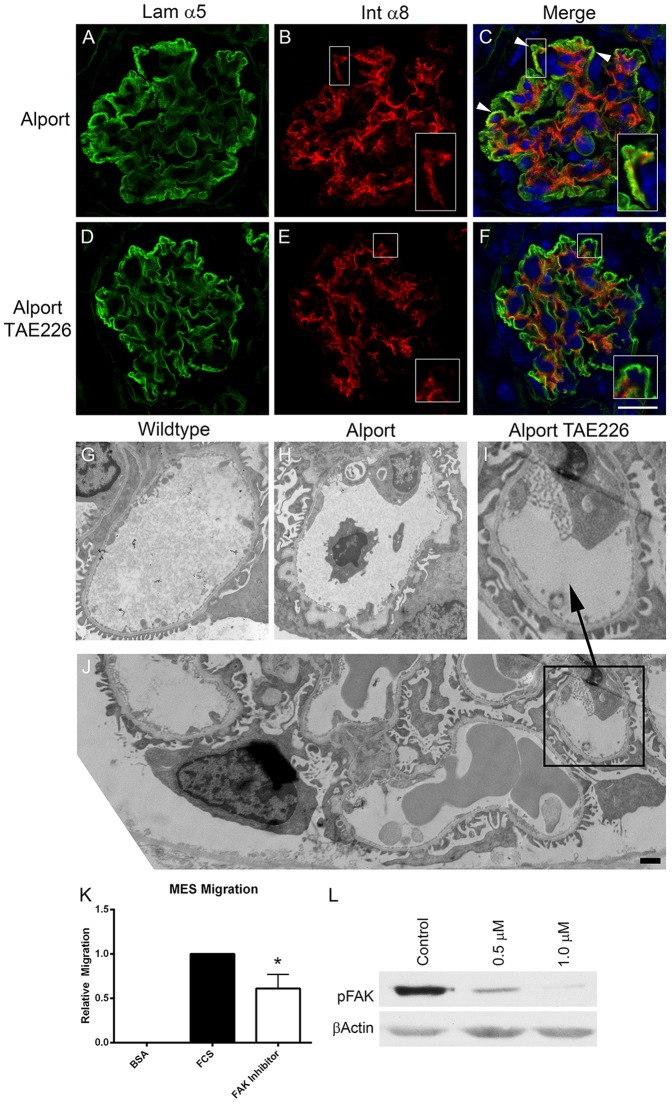
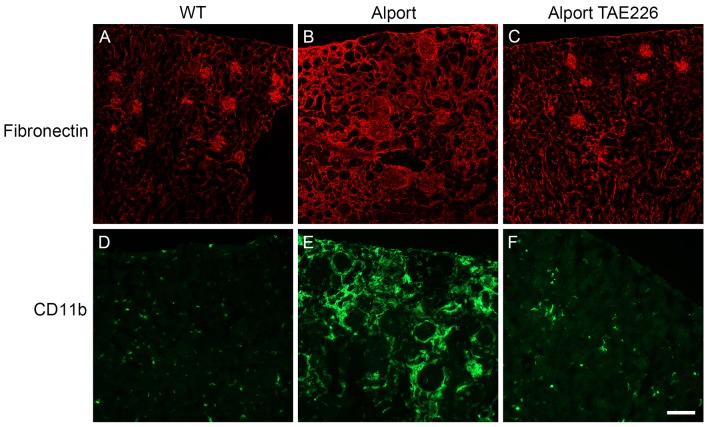
It has been known for some time that laminins containing α1 and α2 chains, which are normally restricted to the mesangial matrix, accumulate in the glomerular basement membranes (GBM) of Alport mice, dogs, and humans. We show that laminins containing the α2 chain, but not those containing the α1 chain activates focal adhesion kinase (FAK) on glomerular podocytes in vitro and in vivo. CD151-null mice, which have weakened podocyte adhesion to the GBM rendering these mice more susceptible to biomechanical strain in the glomerulus, also show progressive accumulation of α2 laminins in the GBM, and podocyte FAK activation. Analysis of glomerular mRNA from both models demonstrates significant induction of MMP-9, MMP-10, MMP-12, MMPs linked to GBM destruction in Alport disease models, as well as the pro-inflammatory cytokine IL-6. SiRNA knockdown of FAK in cultured podocytes significantly reduced expression of MMP-9, MMP-10 and IL-6, but not MMP-12. Treatment of Alport mice with TAE226, a small molecule inhibitor of FAK activation, ameliorated fibrosis and glomerulosclerosis, significantly reduced proteinuria and blood urea nitrogen levels, and partially restored GBM ultrastructure. Glomerular expression of MMP-9, MMP-10 and MMP-12 mRNAs was significantly reduced in TAE226 treated animals. Collectively, this work identifies laminin α2-mediated FAK activation in podocytes as an important early event in Alport glomerular pathogenesis and suggests that FAK inhibitors, if safe formulations can be developed, might be employed as a novel therapeutic approach for treating Alport renal disease in its early stages.
Conflict of interest statement
Figures
References
-
- Kruegel J, Rubel D, Gross O (2013) Alport syndrome—insights from basic and clinical research. Nat Rev Nephrol 9: 170–178. - PubMed
-
- Abrahamson DR, Isom K, Roach E, St John PL (2007) Laminin compensation in collagen alpha3(IV) knockout (Alport) glomeruli contributes to permeability defects. J Am Soc Nephrol 18: 2465–2472. - PubMed
-
- Gunwar S, Ballester F, Noelken ME, Sado Y, Hudson BG (1998) Glomerular basement membrane. Identification of a novel disulfide-cross-linked network of alpha3, alpha4, and alpha5 chains of type IV collagen and its implications for the pathogenesis of Alport syndrome. J Biol Chem 273: 8767–8775. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
