

In vitro analysis of human immunodeficiency virus particle dissociation: gag proteolytic processing influences dissociation kinetics
- PMID: 24915417
- PMCID: PMC4051761
- DOI: 10.1371/journal.pone.0099504
In vitro analysis of human immunodeficiency virus particle dissociation: gag proteolytic processing influences dissociation kinetics
Abstract
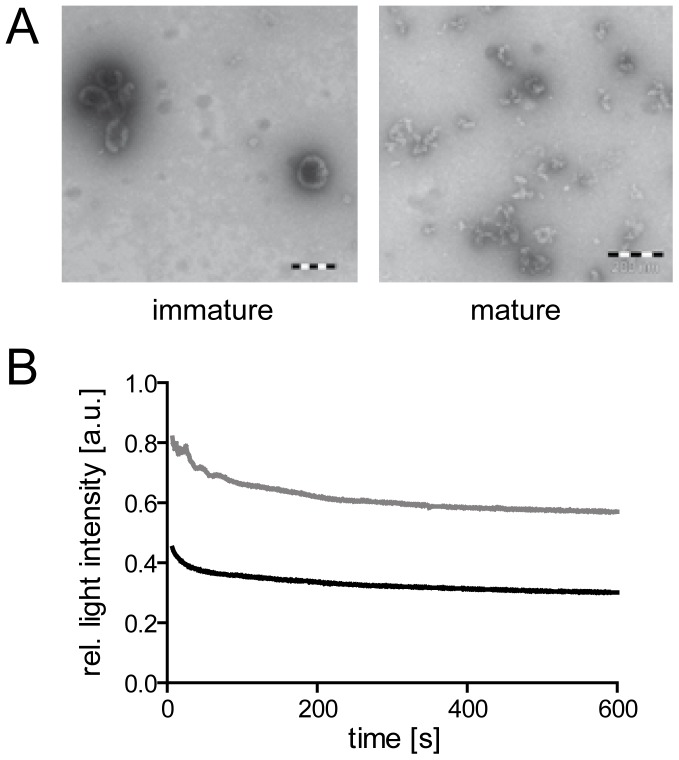
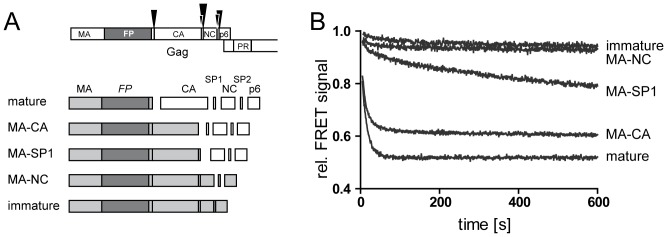
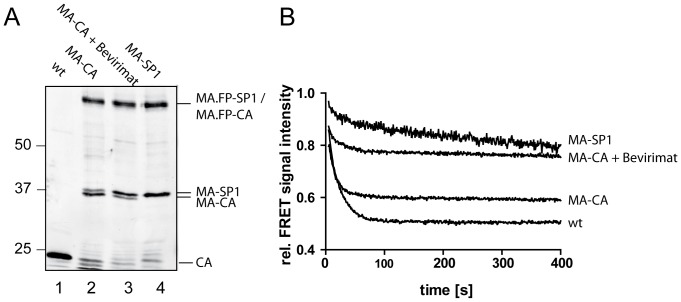
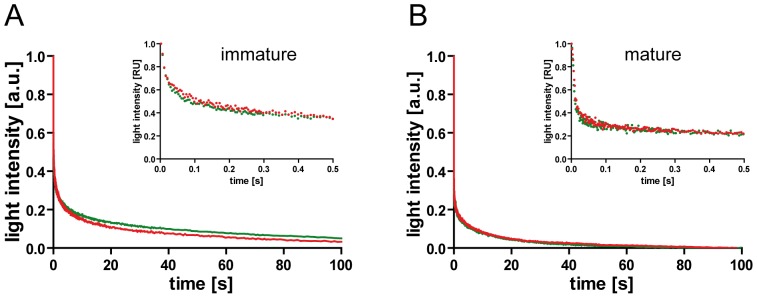
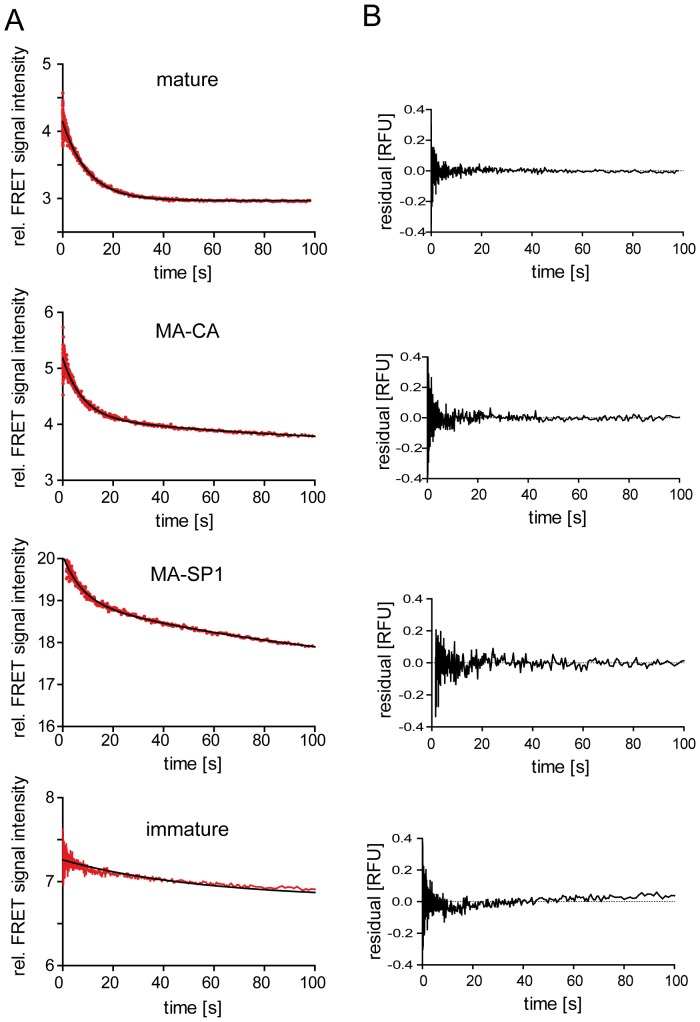
Human immunodeficiency virus particles undergo a step of proteolytic maturation, in which the main structural polyprotein Gag is cleaved into its mature subunits matrix (MA), capsid (CA), nucleocapsid (NC) and p6. Gag proteolytic processing is accompanied by a dramatic structural rearrangement within the virion, which is necessary for virus infectivity and has been proposed to proceed through a sequence of dissociation and reformation of the capsid lattice. Morphological maturation appears to be tightly regulated, with sequential cleavage events and two small spacer peptides within Gag playing important roles by regulating the disassembly of the immature capsid layer and formation of the mature capsid lattice. In order to measure the influence of individual Gag domains on lattice stability, we established Förster's resonance energy transfer (FRET) reporter virions and employed rapid kinetic FRET and light scatter measurements. This approach allowed us to measure dissociation properties of HIV-1 particles assembled in eukaryotic cells containing Gag proteins in different states of proteolytic processing. While the complex dissociation behavior of the particles prevented an assignment of kinetic rate constants to individual dissociation steps, our analyses revealed characteristic differences in the dissociation properties of the MA layer dependent on the presence of additional domains. The most striking effect observed here was a pronounced stabilization of the MA-CA layer mediated by the presence of the 14 amino acid long spacer peptide SP1 at the CA C-terminus, underlining the crucial role of this peptide for the resolution of the immature particle architecture.
Conflict of interest statement
Figures

References
-
- Briggs JA, Kräusslich HG (2011) The molecular architecture of HIV. J Mol Biol 410: 491–500. - PubMed
-
- Helenius A (1992) Unpacking the incoming influenza virus. Cell 69: 577–578. - PubMed
-
- Park J, Morrow CD (1993) Mutations in the protease gene of human immunodeficiency virus type 1 affect release and stability of virus particles. Virology 194: 843–850. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
