

Mechanisms of organelle biogenesis govern stochastic fluctuations in organelle abundance
- PMID: 24916159
- PMCID: PMC4046565
- DOI: 10.7554/eLife.02678
Mechanisms of organelle biogenesis govern stochastic fluctuations in organelle abundance
Erratum in
-
Correction: Mechanisms of organelle biogenesis govern stochastic fluctuations in organelle abundance.Elife. 2015 Nov 24;4:e12522. doi: 10.7554/eLife.12522. Elife. 2015. PMID: 26600005 Free PMC article. No abstract available.
Abstract
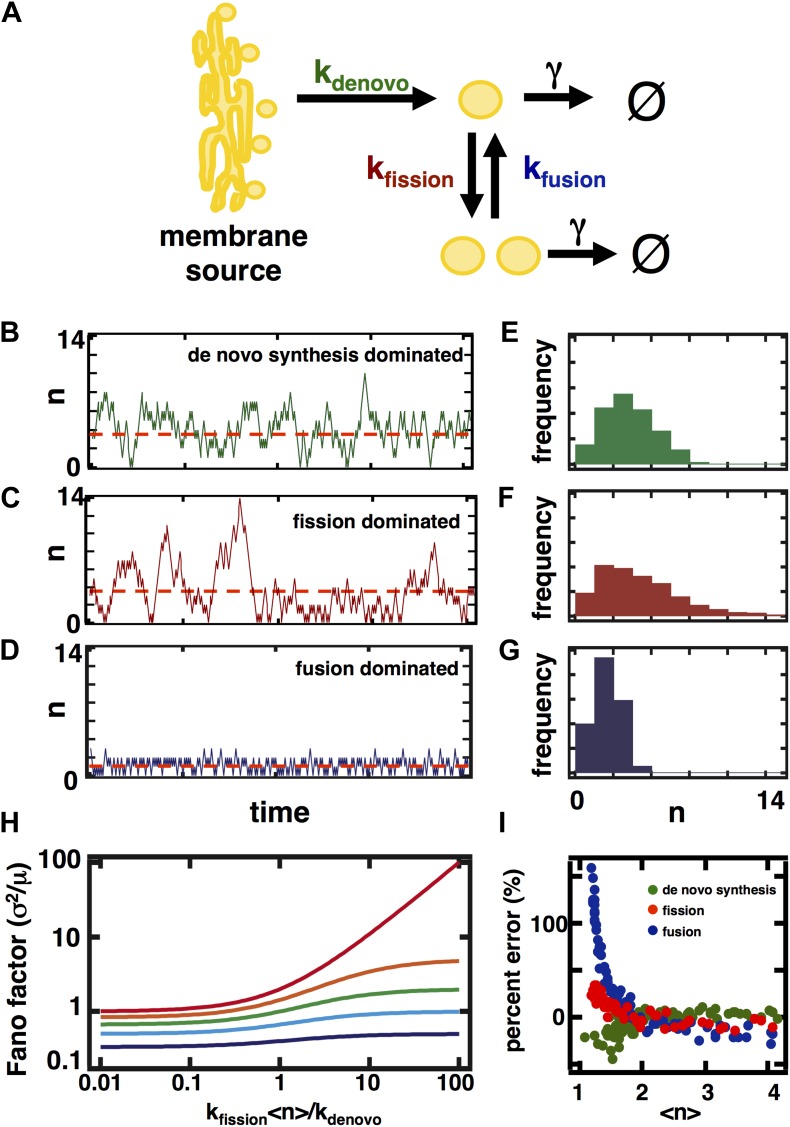
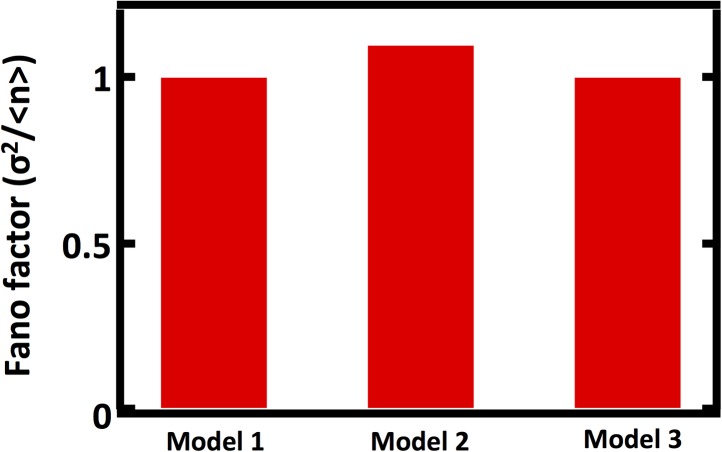
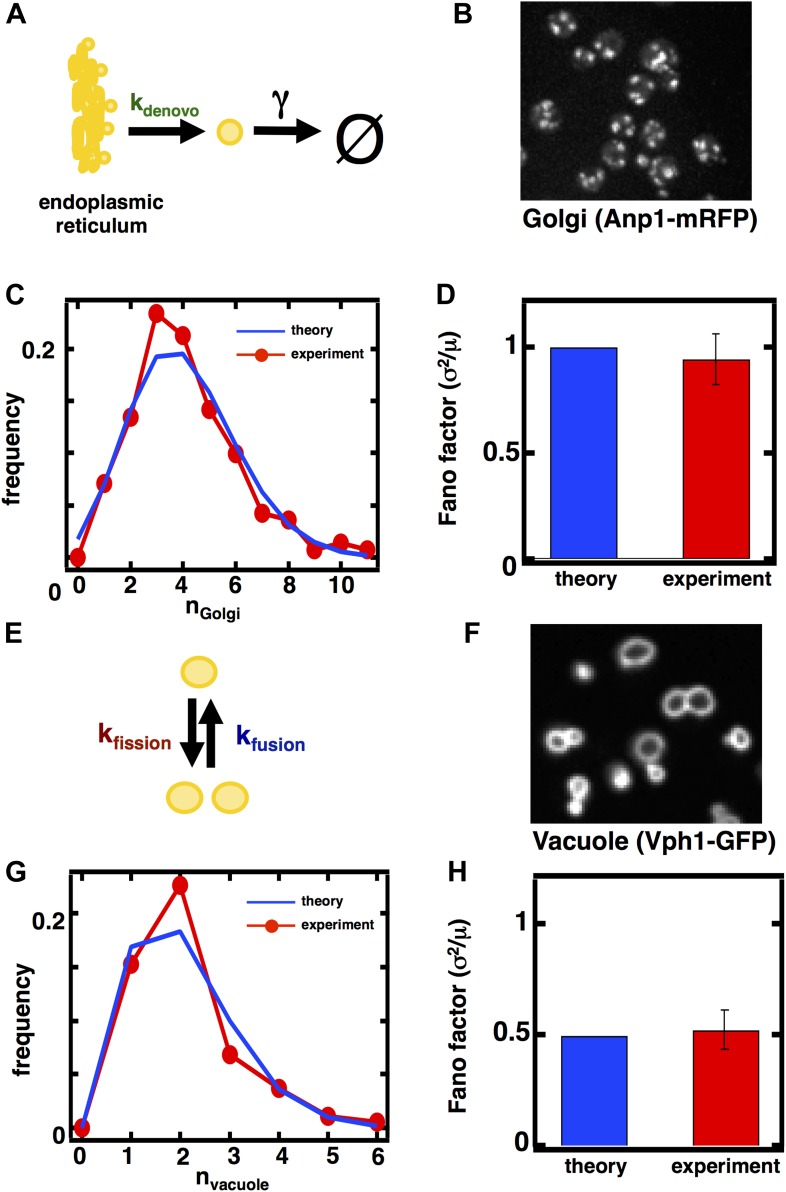
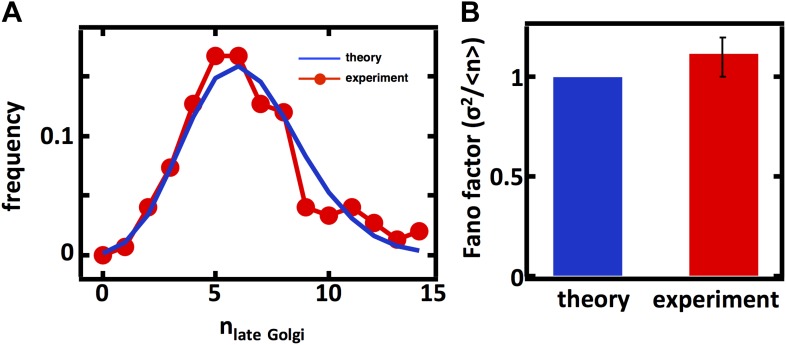
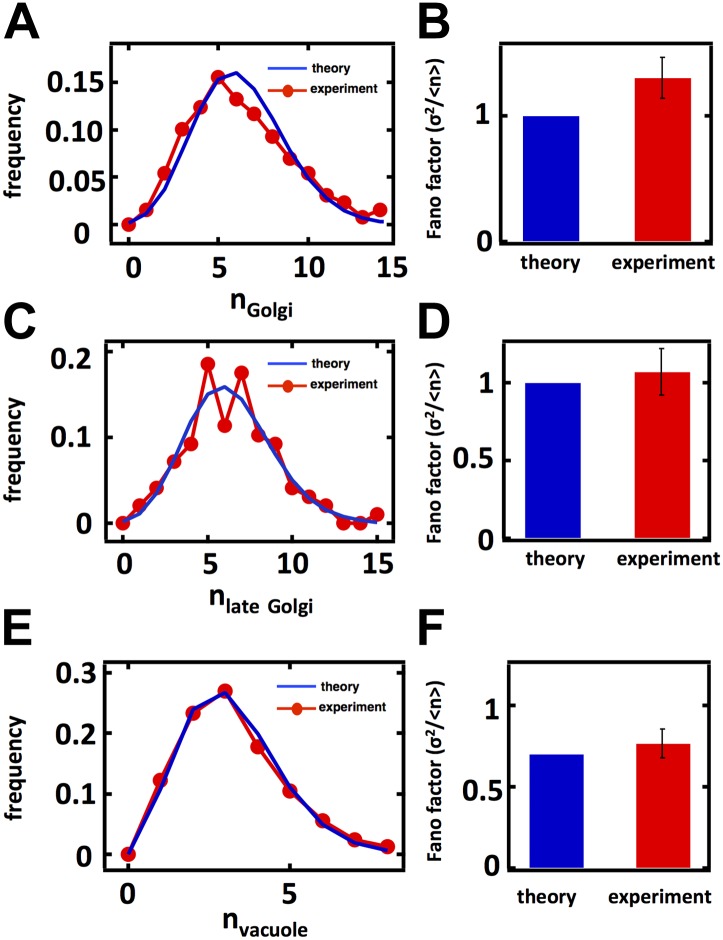
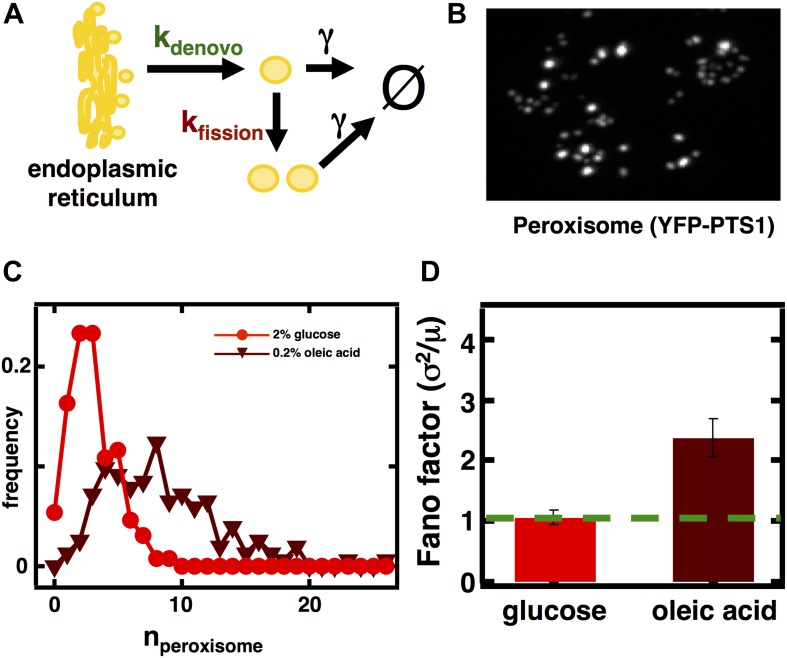
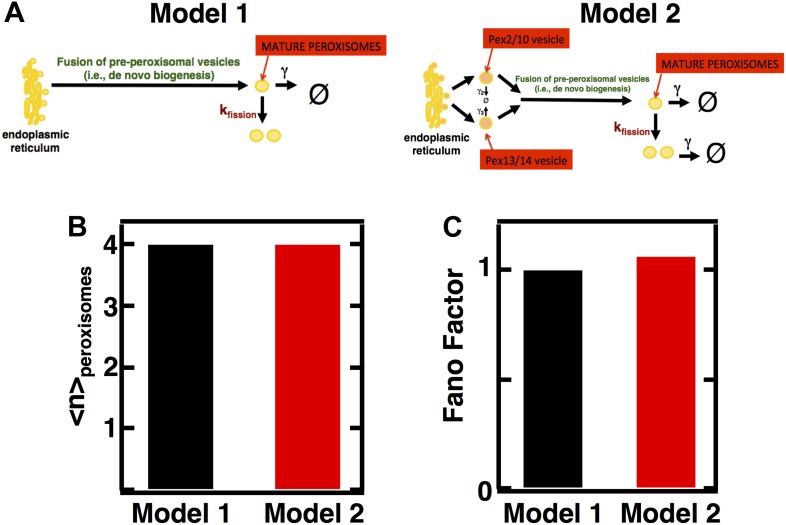
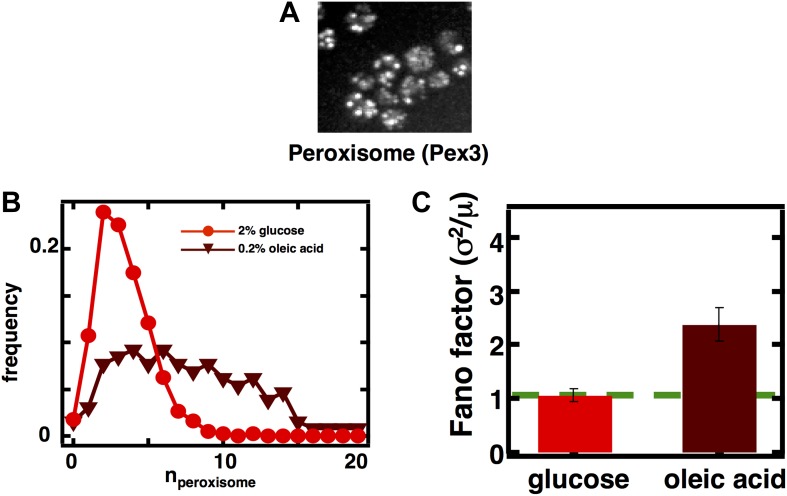
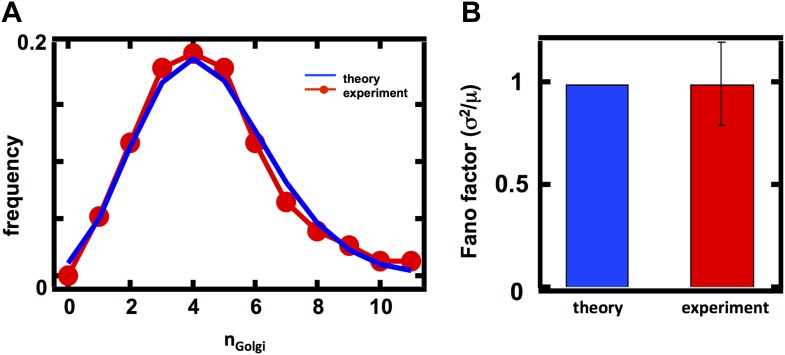
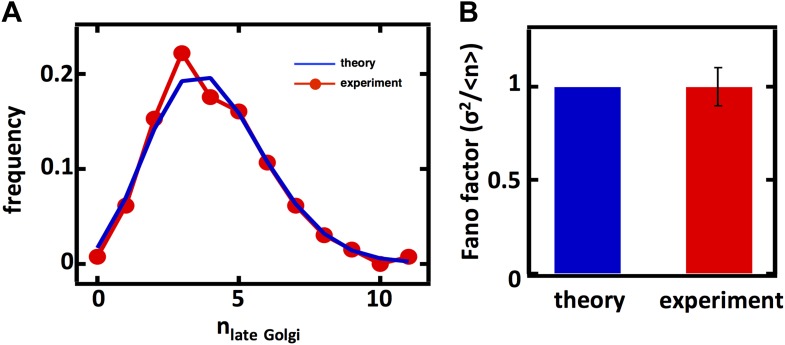
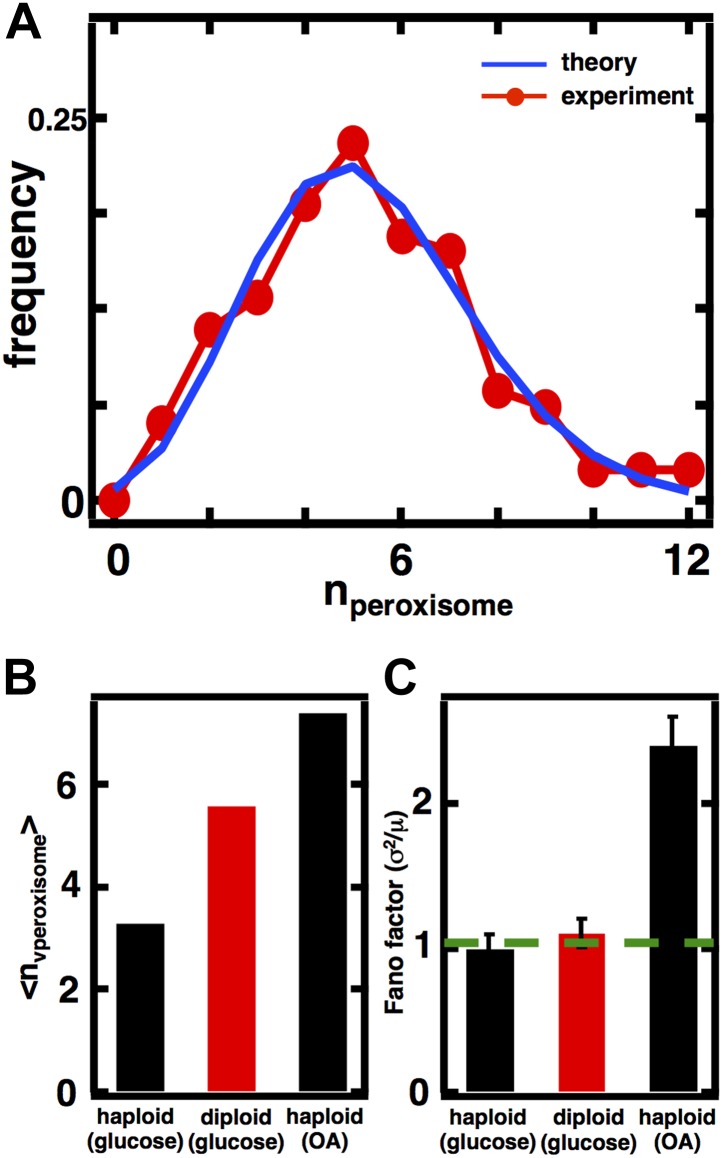
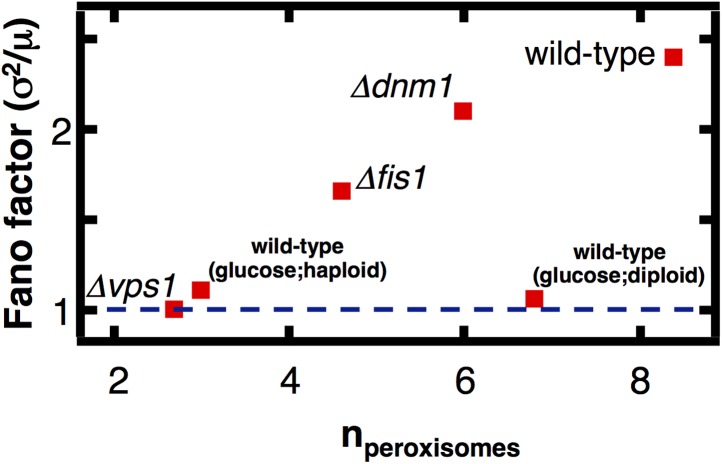
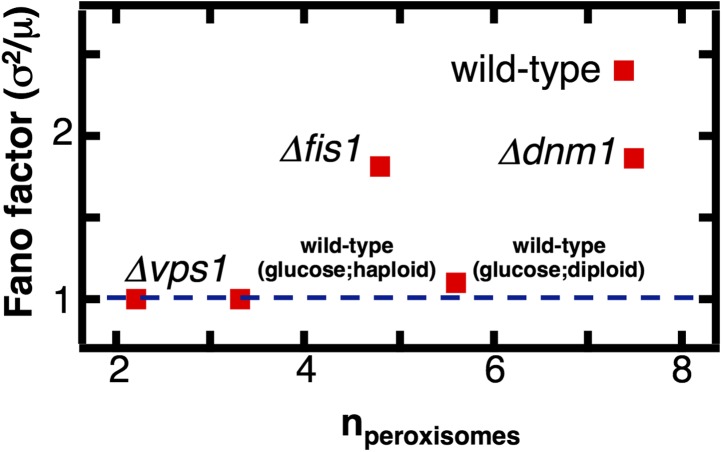
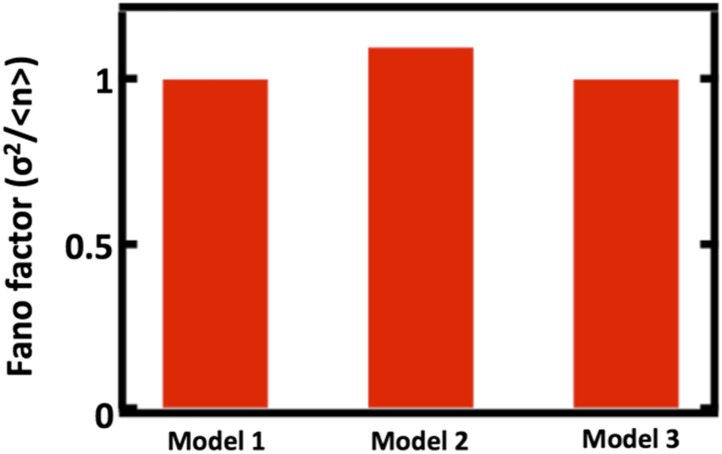
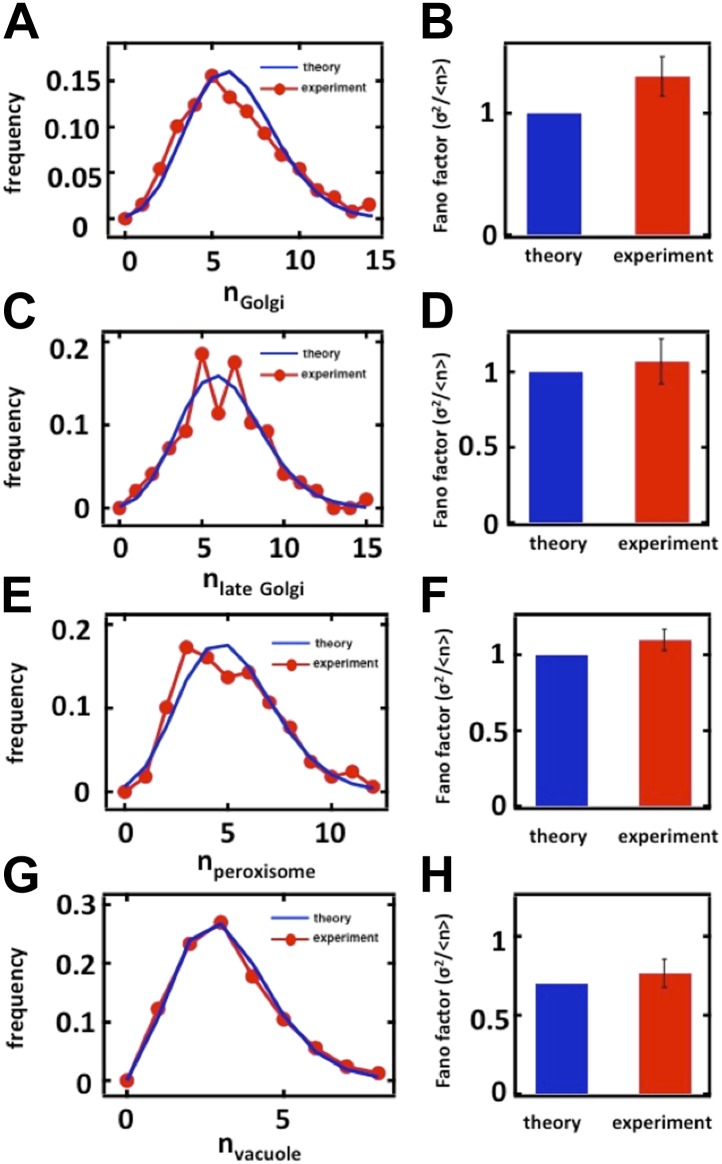
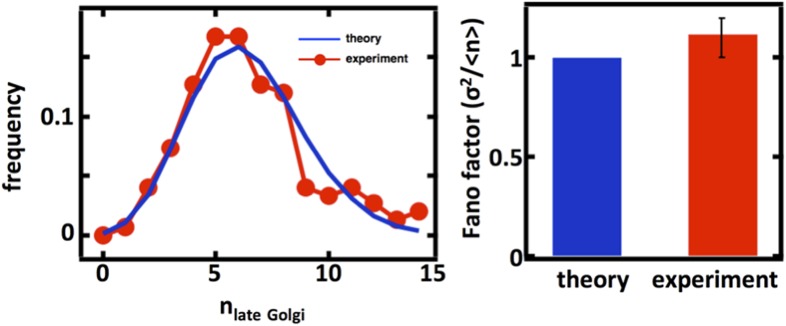
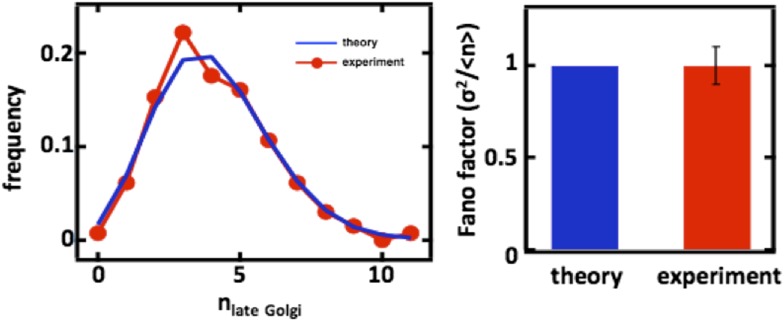
Fluctuations in organelle abundance can profoundly limit the precision of cell biological processes from secretion to metabolism. We modeled the dynamics of organelle biogenesis and predicted that organelle abundance fluctuations depend strongly on the specific mechanisms that increase or decrease the number of a given organelle. Our model exactly predicts the size of experimentally measured Golgi apparatus and vacuole abundance fluctuations, suggesting that cells tolerate the maximum level of variability generated by the Golgi and vacuole biogenesis pathways. We observe large increases in peroxisome abundance fluctuations when cells are transferred from glucose-rich to fatty acid-rich environments. These increased fluctuations are significantly diminished in mutants lacking peroxisome fission factors, leading us to infer that peroxisome biogenesis switches from de novo synthesis to primarily fission. Our work provides a general framework for exploring stochastic organelle biogenesis and using fluctuations to quantitatively unravel the biophysical pathways that control the abundance of subcellular structures.DOI: http://dx.doi.org/10.7554/eLife.02678.001.
Keywords: cell biology; fluctuations; mathematical modeling; organelles; systems biology.
Copyright © 2014, Mukherji and O'Shea.
Conflict of interest statement
EKO: Erin O'Shea is Chief Scientific Officer and a Vice President at the Howard Hughes Medical Institute, one of the three founding funders of
The other author declares that no competing interests exist.
Figures
References
-
- Bhave M, Papanikou E, Iyer P, Pandya K, Jain BK, Ganguly A, Sharma C, Pawar K, Austin J, II, Day KJ, Rossanese OW, Glick BS, Bhattacharyya D. 2014. Golgi enlargement in Arf-depleted yeast cells is due to altered dynamics of cisternal maturation. Journal of Cell Science 127:250–257. doi: 10.1242/jcs.140996 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
