

Fructose 1-phosphate is the one and only physiological effector of the Cra (FruR) regulator of Pseudomonas putida
- PMID: 24918052
- PMCID: PMC4050194
- DOI: 10.1016/j.fob.2014.03.013
Fructose 1-phosphate is the one and only physiological effector of the Cra (FruR) regulator of Pseudomonas putida
Abstract
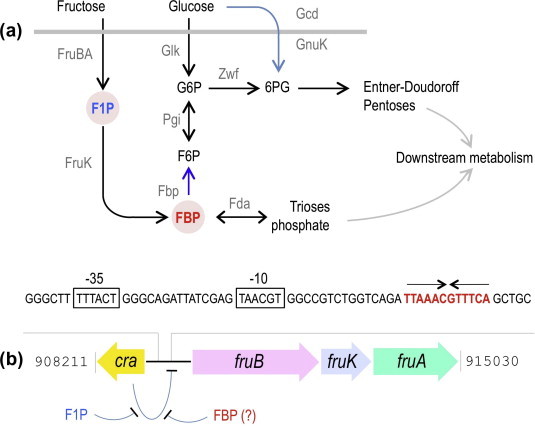
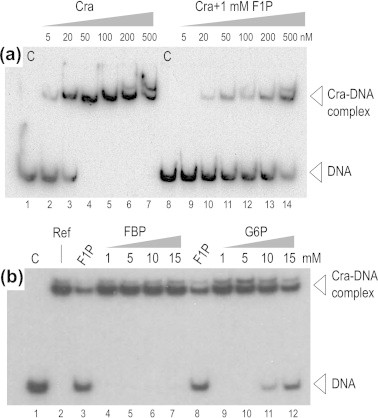
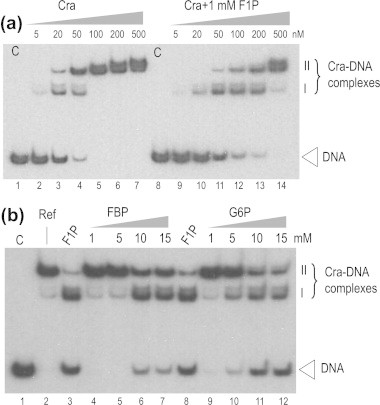
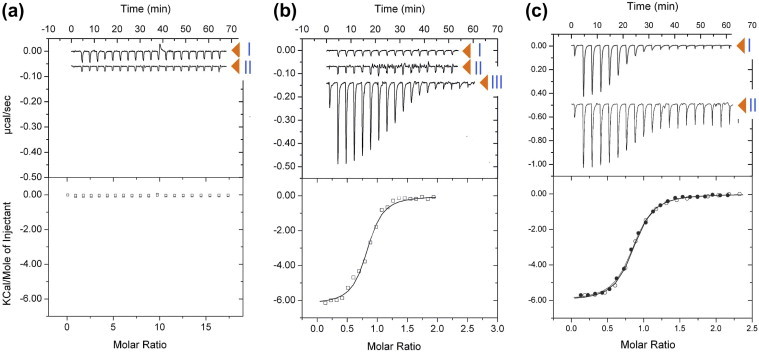
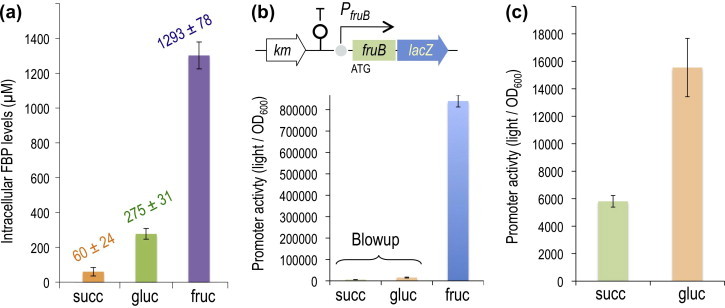
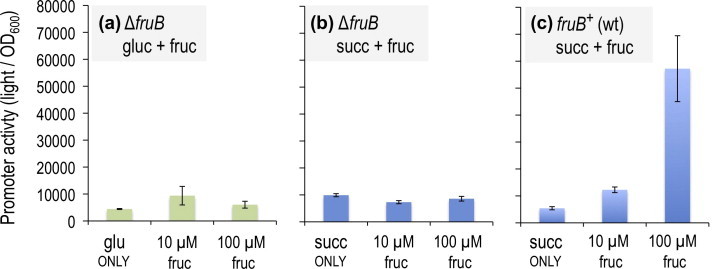
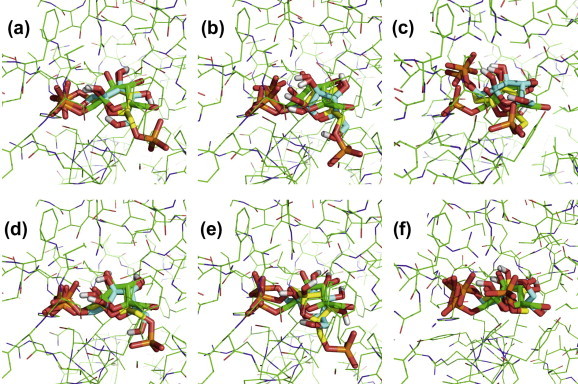
Fructose-1-phosphate (F1P) is the preferred effector of the catabolite repressor/activator (Cra) protein of the soil bacterium Pseudomonas putida but its ability to bind other metabolic intermediates in vivo is unclear. The Cra protein of this microorganism (Cra(PP)) was submitted to mobility shift assays with target DNA sequences (the PfruB promoter) and candidate effectors fructose-1,6-bisphosphate (FBP), glucose 6-phosphate (G6P), and fructose-6-phosphate (F6P). 1 mM F1P was sufficient to release most of the Cra protein from its operators but more than 10 mM of FBP or G6P was required to free the same complex. However, isothermal titration microcalorimetry failed to expose any specific interaction between Cra(PP) and FBP or G6P. To solve this paradox, transcriptional activity of a PfruB-lacZ fusion was measured in wild-type and ΔfruB cells growing on substrates that change the intracellular concentrations of F1P and FBP. The data indicated that PfruB activity was stimulated by fructose but not by glucose or succinate. This suggested that Cra(PP) represses expression in vivo of the cognate fruBKA operon in a fashion dependent just on F1P, ruling out any other physiological effector. Molecular docking and dynamic simulations of the Cra-agonist interaction indicated that both metabolites can bind the repressor, but the breach in the relative affinity of Cra(PP) for F1P vs FBP is three orders of magnitude larger than the equivalent distance in the Escherichia coli protein. This assigns the Cra protein of P. putida the sole role of transducing the presence of fructose in the medium into a variety of direct and indirect physiological responses.
Keywords: Cra; Cra, catabolic repression/activation protein; F1P, fructose-1-phosphate; F6P, fructose-6-phosphate; FBP, fructose-1,6-bisphosphate; FruR; Fructose 1-phosphate; Fructose operon; G6P, glucose 6-phosphate; ITC, isothermal calorimetry; Pseudomonas putida.
Figures
References
-
- Ramseier T.M. Cra and the control of carbon flux via metabolic pathways. Res. Microbiol. 1996;147:489–493. - PubMed
-
- Ow D.S., Lee R.M., Nissom P.M., Philp R., Oh S.K., Yap M.G. Inactivating FruR global regulator in plasmid-bearing Escherichia coli alters metabolic gene expression and improves growth rate. J. Biotechnol. 2007;131:261–269. - PubMed
-
- Sarkar D., Siddiquee K.A., Arauzo-Bravo M.J., Oba T., Shimizu K. Effect of cra gene knockout together with edd and iclR genes knockout on the metabolism in Escherichia coli. Arch. Microbiol. 2008;190:559–571. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
