

The Nogo receptor NgR1 mediates infection by mammalian reovirus
- PMID: 24922571
- PMCID: PMC4100558
- DOI: 10.1016/j.chom.2014.05.010
The Nogo receptor NgR1 mediates infection by mammalian reovirus
Abstract
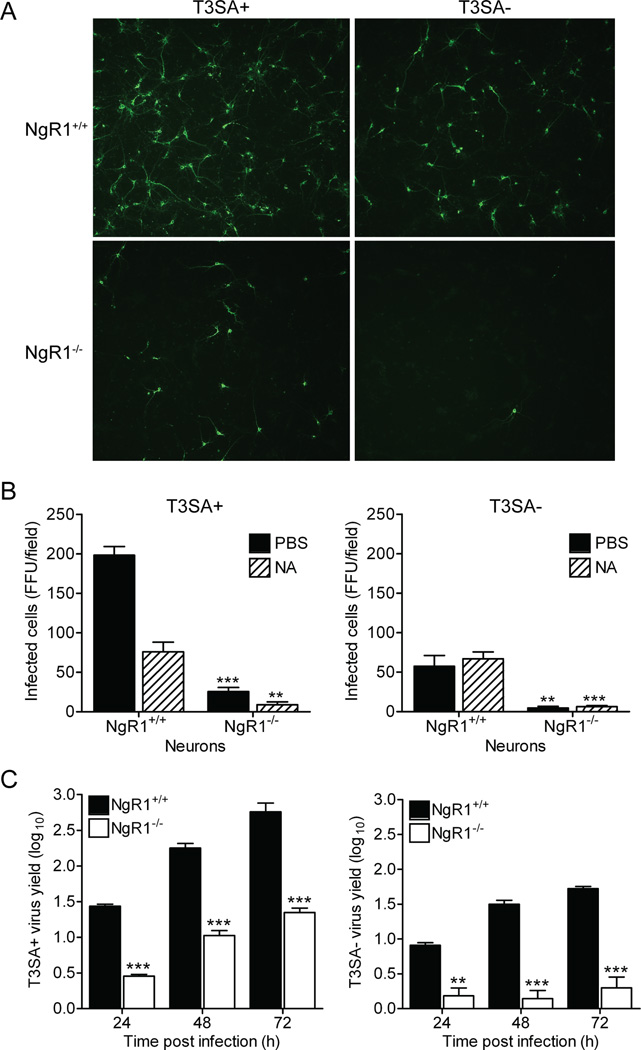
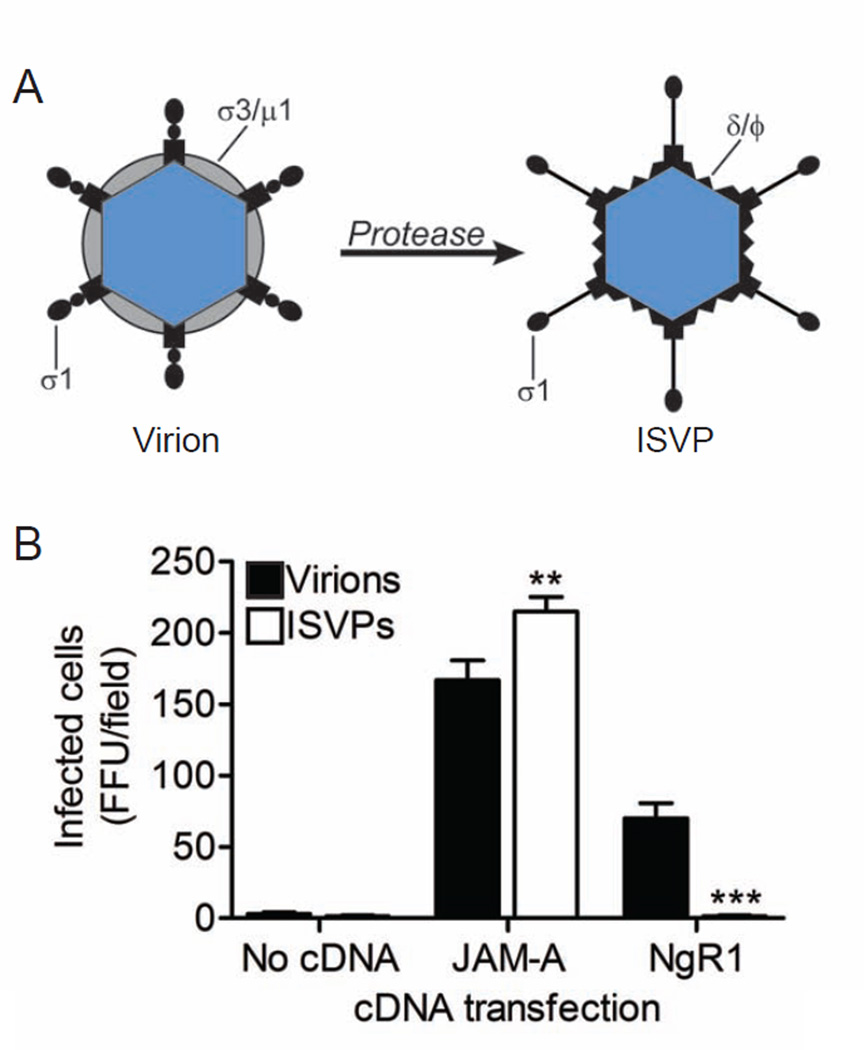
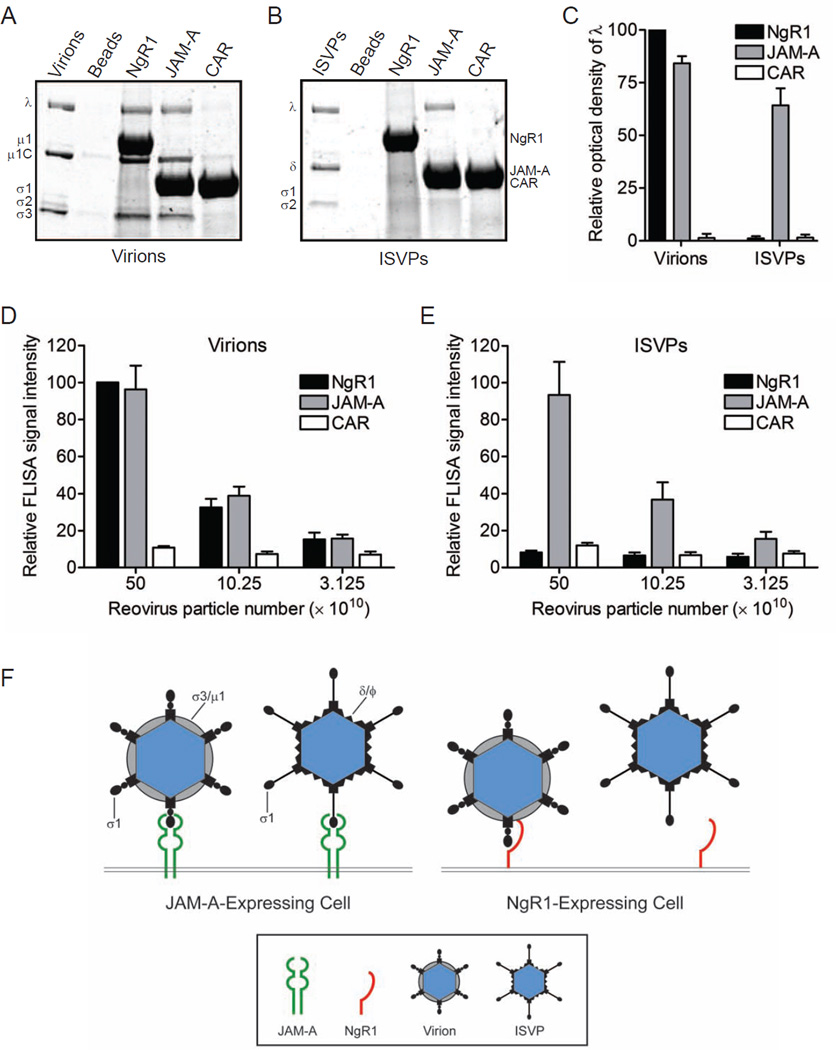
Neurotropic viruses, including mammalian reovirus, must disseminate from an initial site of replication to the central nervous system (CNS), often binding multiple receptors to facilitate systemic spread. Reovirus engages junctional adhesion molecule A (JAM-A) to disseminate hematogenously. However, JAM-A is dispensable for reovirus replication in the CNS. We demonstrate that reovirus binds Nogo receptor NgR1, a leucine-rich repeat protein expressed in the CNS, to infect neurons. Expression of NgR1 confers reovirus binding and infection of nonsusceptible cells. Incubating reovirus virions with soluble NgR1 neutralizes infectivity. Blocking NgR1 on transfected cells or primary cortical neurons abrogates reovirus infection. Concordantly, reovirus infection is ablated in primary cortical neurons derived from NgR1 null mice. Reovirus virions bind to soluble JAM-A and NgR1, while infectious disassembly intermediates (ISVPs) bind only to JAM-A. These results suggest that reovirus uses different capsid components to bind distinct cell-surface molecules, engaging independent receptors to facilitate spread and tropism.
Copyright © 2014 Elsevier Inc. All rights reserved.
Figures
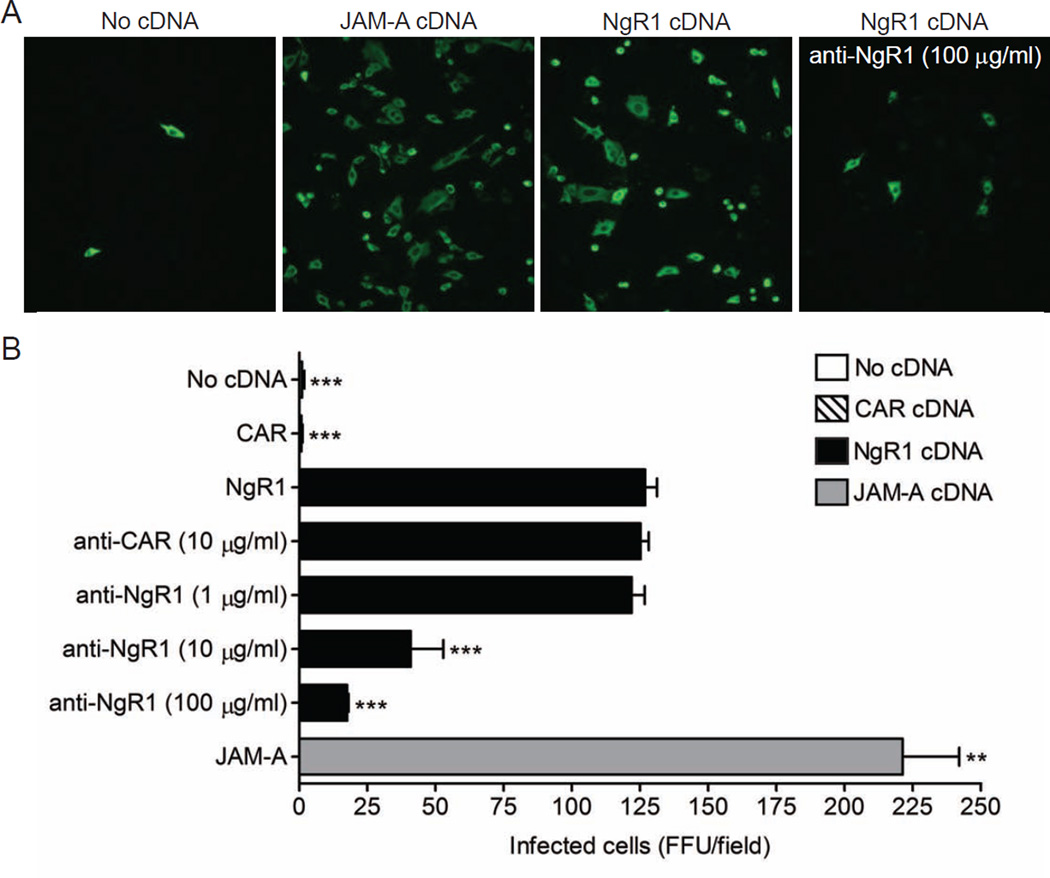
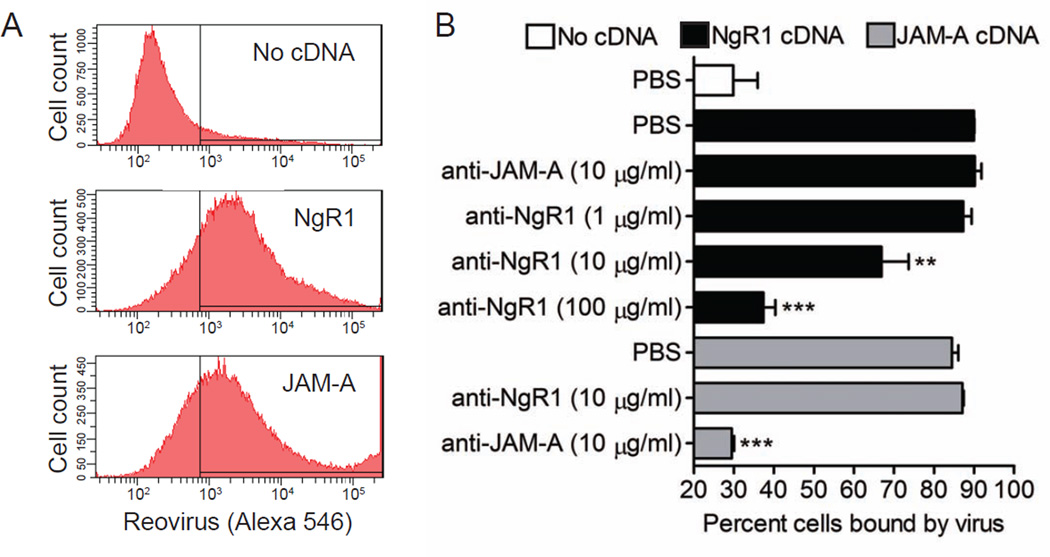
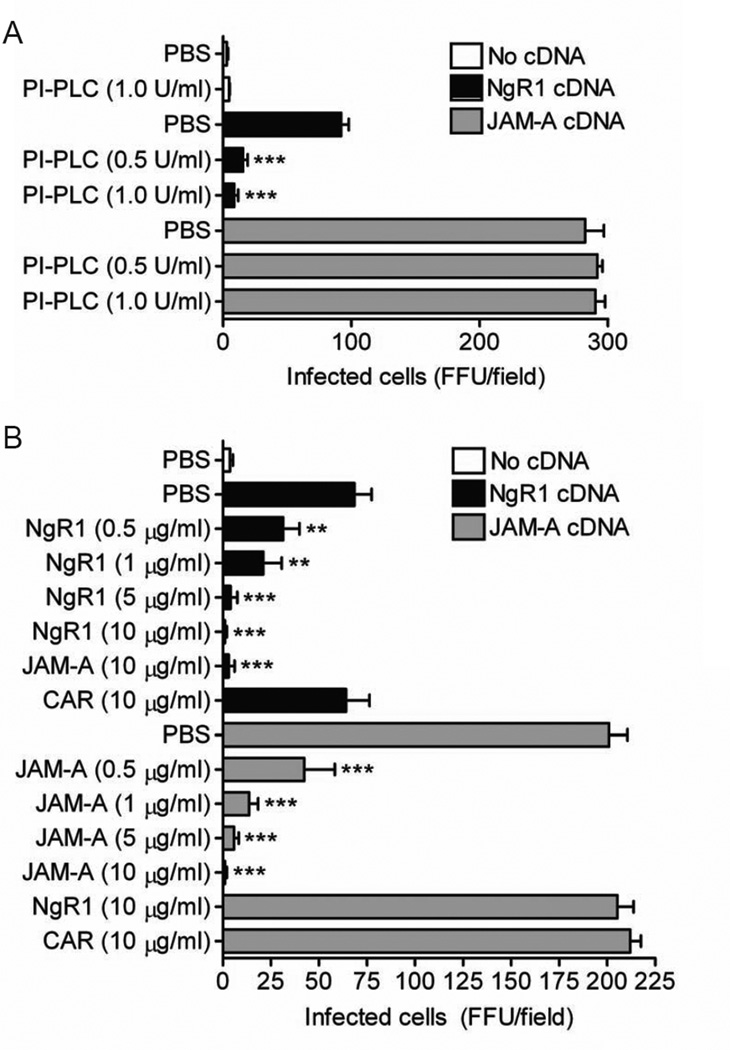
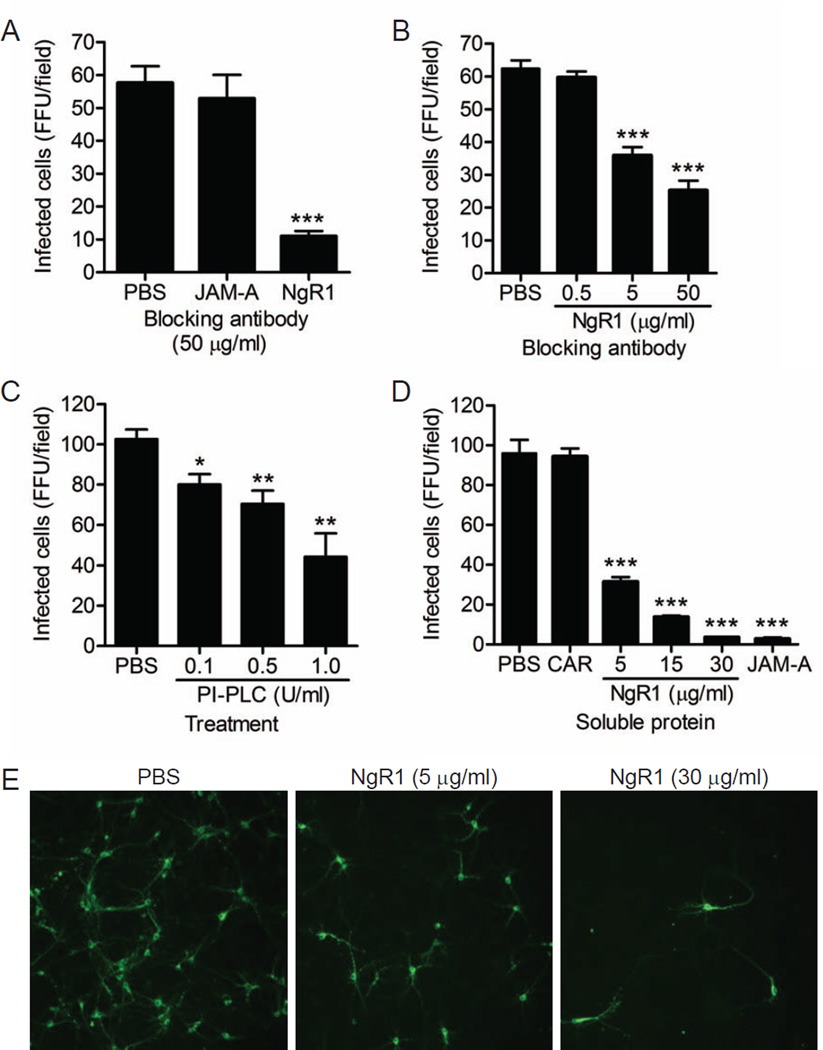
Comment in
-
Viral pathogenesis: Neural receptor for reovirus revealed.Nat Rev Microbiol. 2014 Aug;12(8):532. doi: 10.1038/nrmicro3307. Epub 2014 Jun 23. Nat Rev Microbiol. 2014. PMID: 24954379 No abstract available.
References
-
- Barton ES, Connolly JL, Forrest JC, Chappell JD, Dermody TS. Utilization of sialic acid as a coreceptor enhances reovirus attachment by multistep adhesion strengthening. J. Biol. Chem. 2001a;276:2200–2211. - PubMed
-
- Barton ES, Forrest JC, Connolly JL, Chappell JD, Liu Y, Schnell F, Nusrat A, Parkos CA, Dermody TS. Junction adhesion molecule is a receptor for reovirus. Cell. 2001b;104:441–451. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
