

p62/SQSTM1 plays a protective role in oxidative injury of steatotic liver in a mouse hepatectomy model
- PMID: 24925527
- PMCID: PMC4245881
- DOI: 10.1089/ars.2013.5391
p62/SQSTM1 plays a protective role in oxidative injury of steatotic liver in a mouse hepatectomy model
Abstract
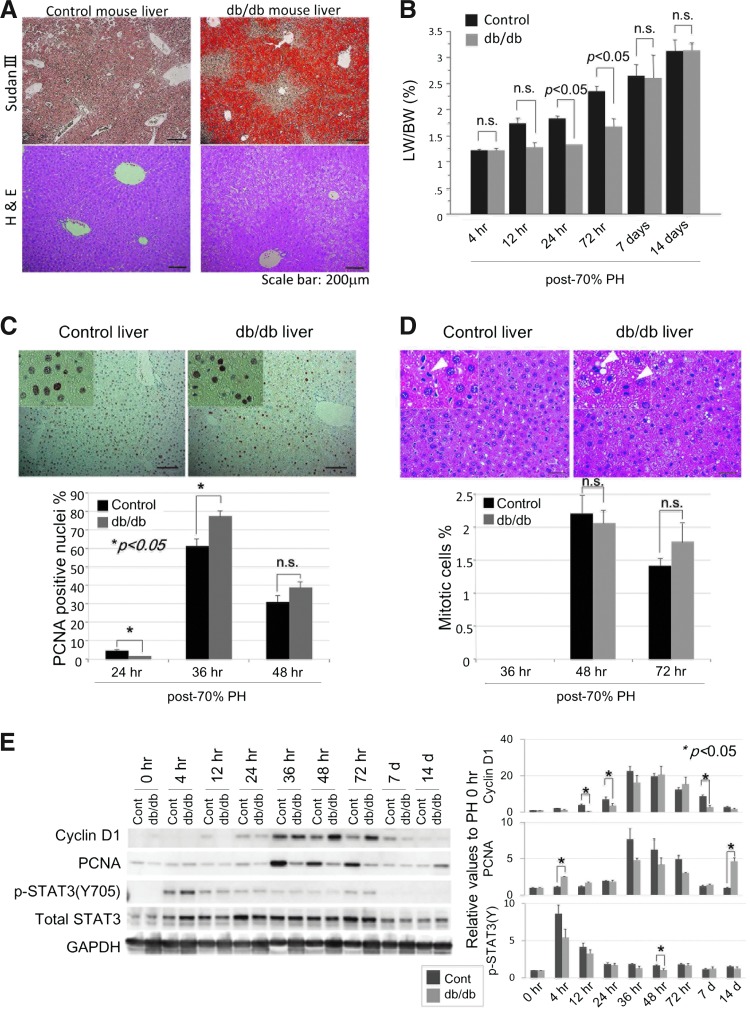
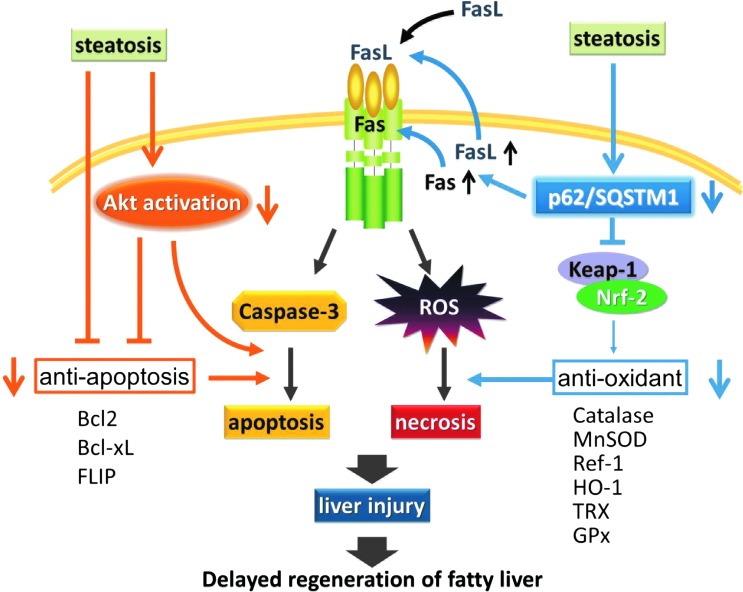
Aims: Liver injury and regeneration involve complicated processes and are affected by various physio-pathological factors. We investigated the mechanisms of steatosis-associated liver injury and delayed regeneration in a mouse model of partial hepatectomy.
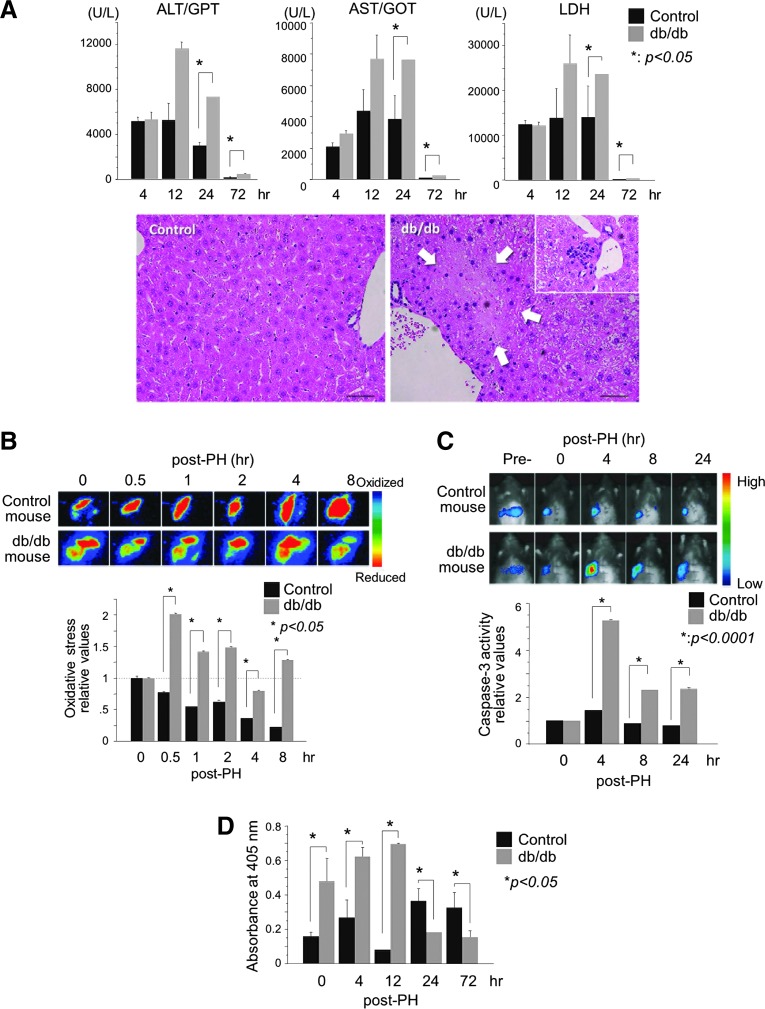
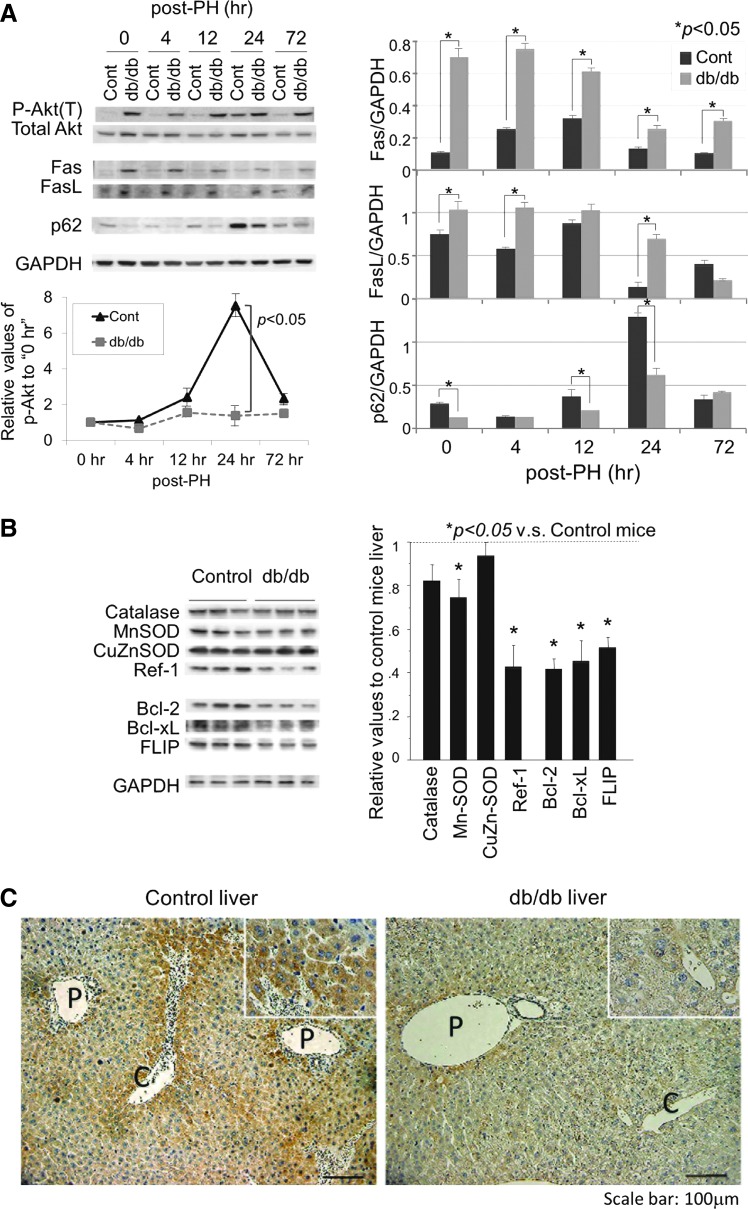
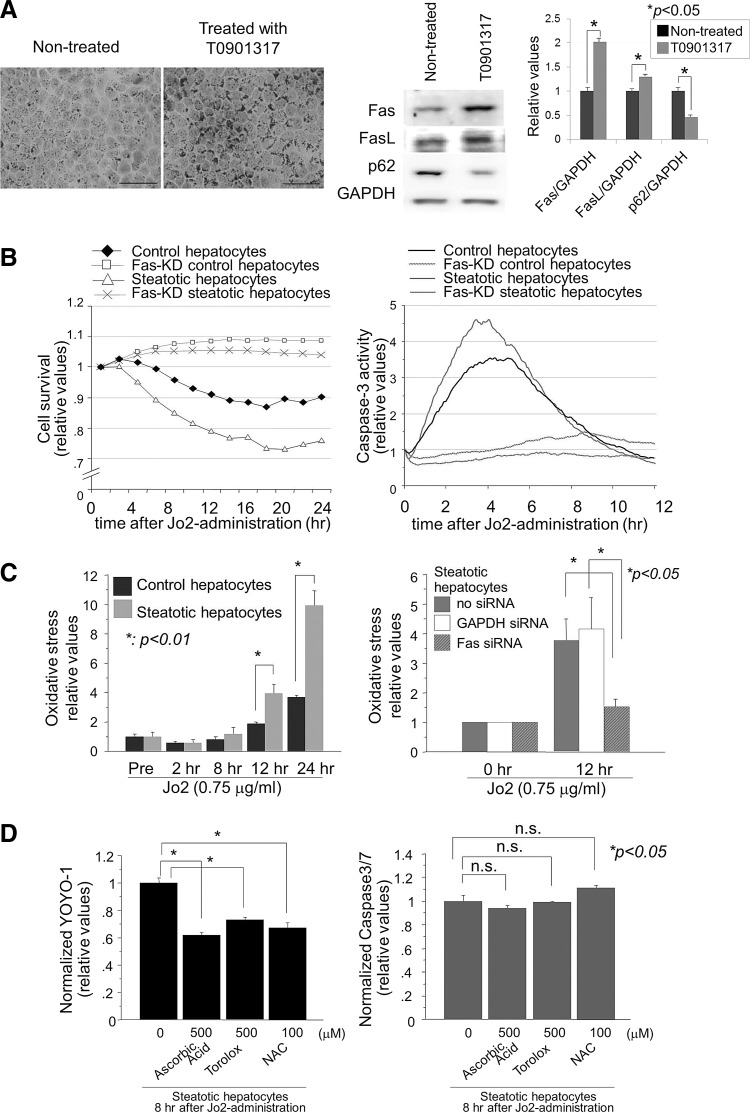
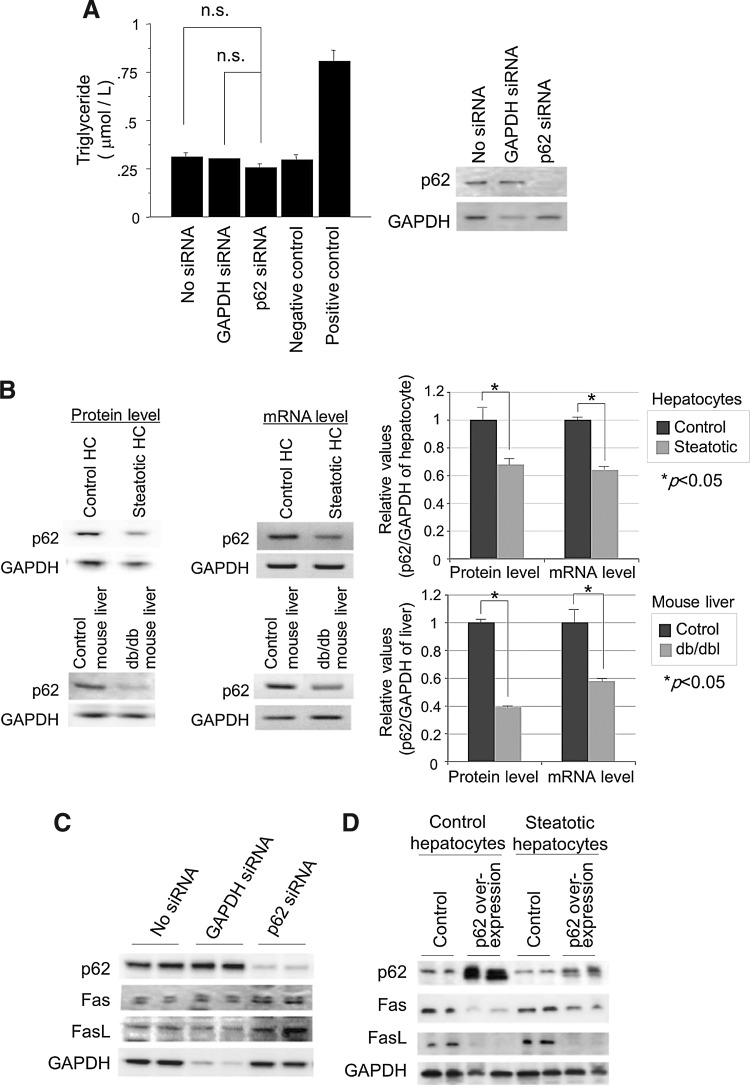
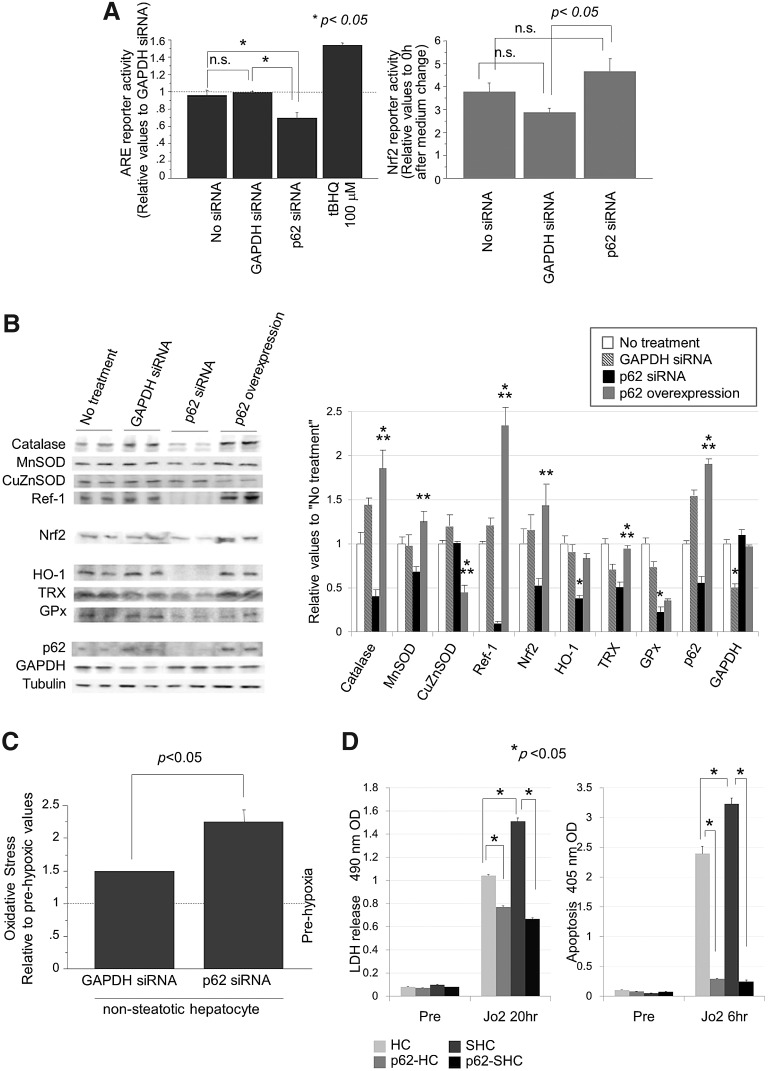
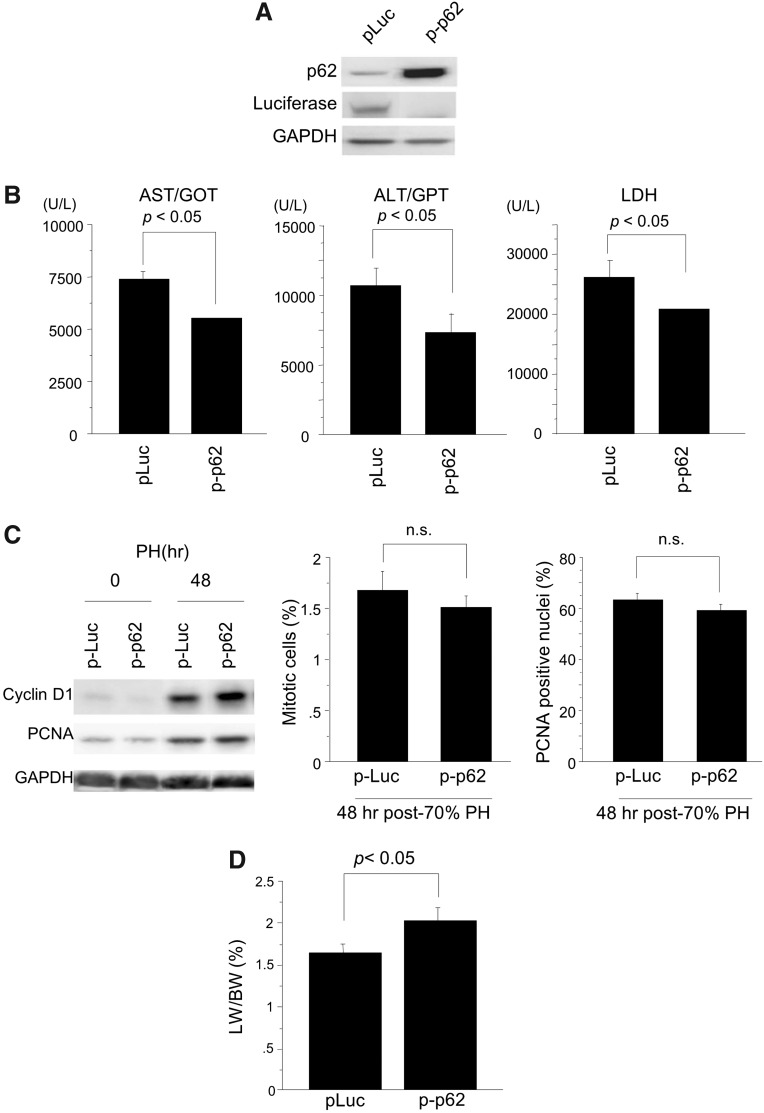
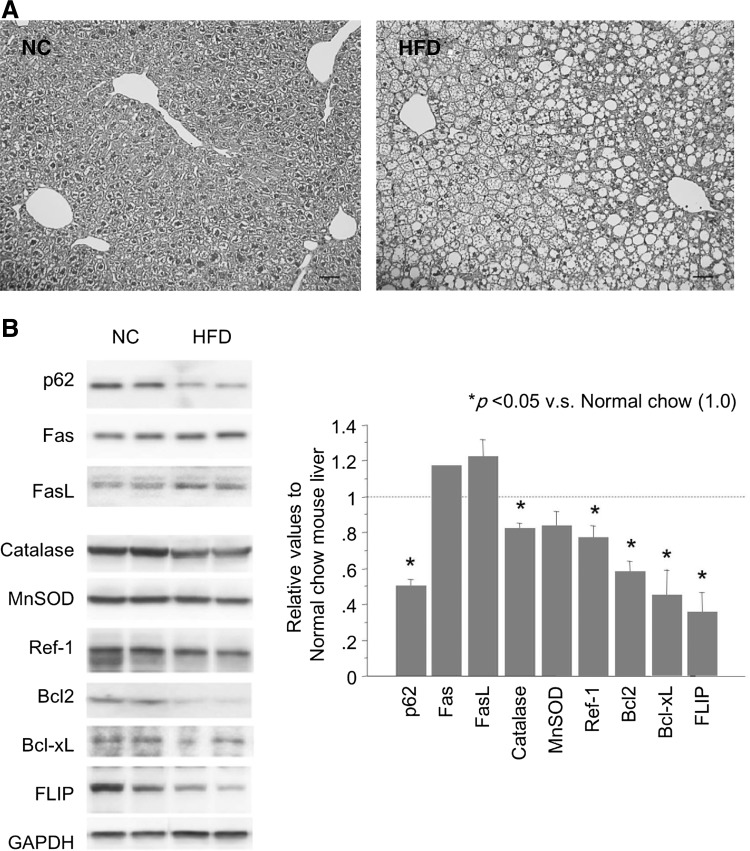
Results: Initial regeneration of the steatotic liver was significantly delayed after hepatectomy. Although hepatocyte proliferation was not significantly suppressed, severe liver injury with oxidative stress (OS) occurred immediately after hepatectomy in the steatotic liver. Fas-ligand (FasL)/Fas expression was upregulated in the steatotic liver, whereas the expression of antioxidant and anti-apoptotic molecules (catalase/MnSOD/Ref-1 and Bcl-2/Bcl-xL/FLIP, respectively) and p62/SQSTM1, a steatosis-associated protein, was downregulated. Interestingly, pro-survival Akt was not activated in response to hepatectomy, although it was sufficiently expressed even before hepatectomy. Suppression of p62/SQSTM1 increased FasL/Fas expression and reduced nuclear factor erythroid 2-related factor-2 (Nrf-2)-dependent antioxidant response elements activity and antioxidant responses in steatotic and nonsteatotic hepatocytes. Exogenously added FasL induced severe cellular OS and necrosis/apoptosis in steatotic hepatocytes, with only the necrosis being inhibited by pretreatment with antioxidants, suggesting that FasL/Fas-induced OS mainly leads to necrosis. Furthermore, p62/SQSTM1 re-expression in the steatotic liver markedly reduced liver injury and improved liver regeneration.
Innovation: This study is the first which demonstrates that reduced expression of p62/SQSTM1 plays a crucial role in posthepatectomy acute injury and delayed regeneration of steatotic liver, mainly via redox-dependent mechanisms.
Conclusion: In the steatotic liver, reduced expression of p62/SQSTM1 induced FasL/Fas overexpression and suppressed antioxidant genes, mainly through Nrf-2 inactivation, which, along with the hypo-responsiveness of Akt, caused posthepatectomy necrotic/apoptotic liver injury and delayed regeneration, both mainly via a redox-dependent mechanism.
Figures
References
-
- Aoyama T, Ikejima K, Kon K, Okumura K, Arai K, and Watanabe S. Pioglitazone promotes survival and prevents hepatic regeneration failure after partial hepatectomy in obese and diabetic KK-A(y) mice. Hepatology 49: 1636–1644, 2009 - PubMed
-
- Caraceni P, Domenicali M, Vendemiale G, Grattagliano I, Pertosa A, Nardo B, Morselli-Labate AM, Trevisani F, Palasciano G, Altomare E, and Bernardi M. The reduced tolerance of rat steatotic liver to ischemia reperfusion is associated with mitochondrial oxidative injury. J Surg Res 124: 160–168, 2005 - PubMed
-
- Cha JY. and Repa JJ. The liver X receptor (LXR) and hepatic lipogenesis. The carbohydrate-response element-binding protein is a target gene of LXR. J Biol Chem 282: 743–751, 2007 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
