

Allele-specific silencing of mutant huntingtin in rodent brain and human stem cells
- PMID: 24926995
- PMCID: PMC4057216
- DOI: 10.1371/journal.pone.0099341
Allele-specific silencing of mutant huntingtin in rodent brain and human stem cells
Abstract
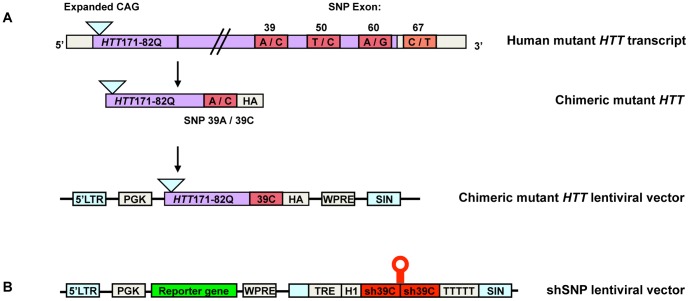
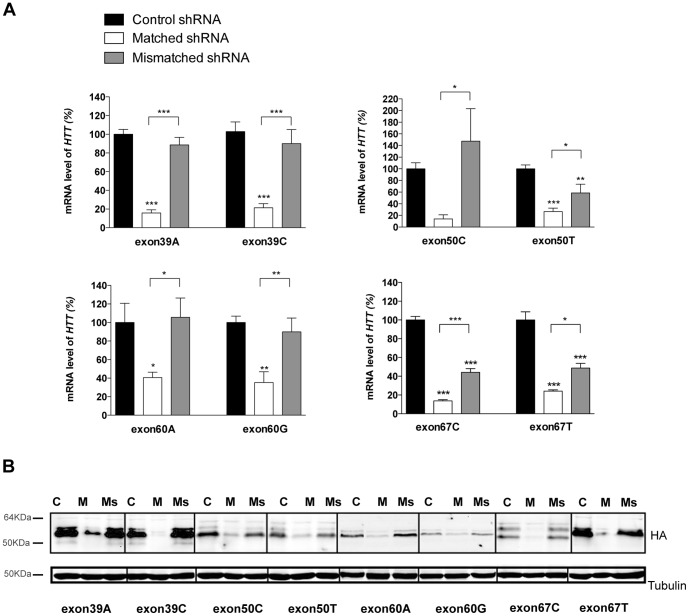
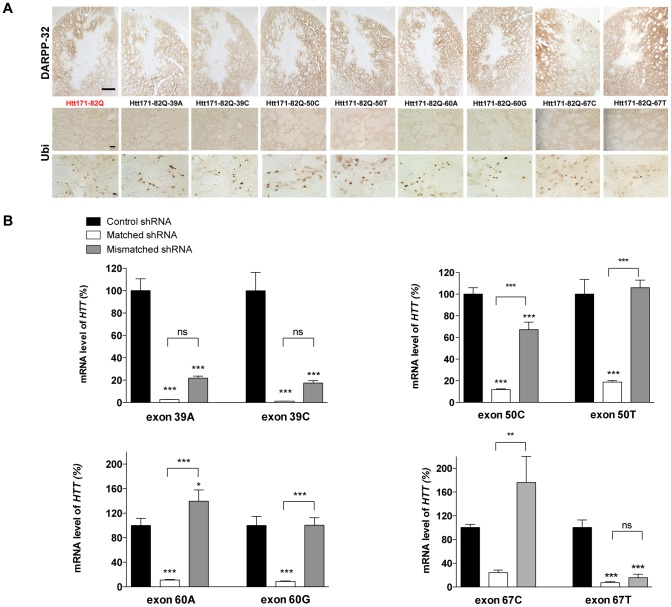
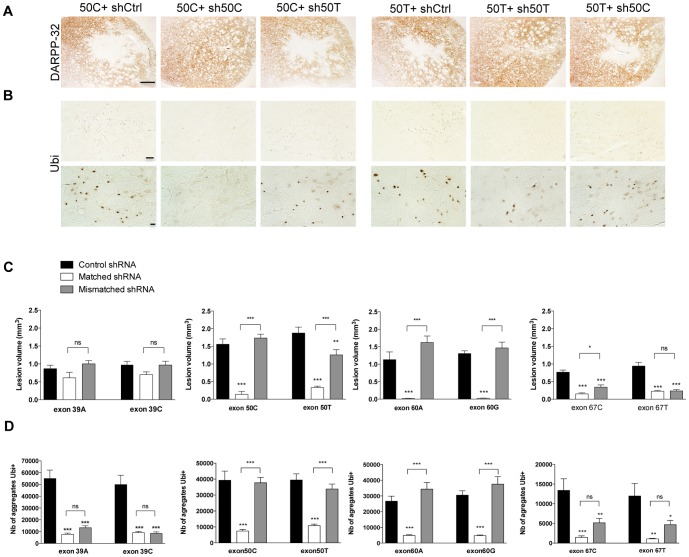
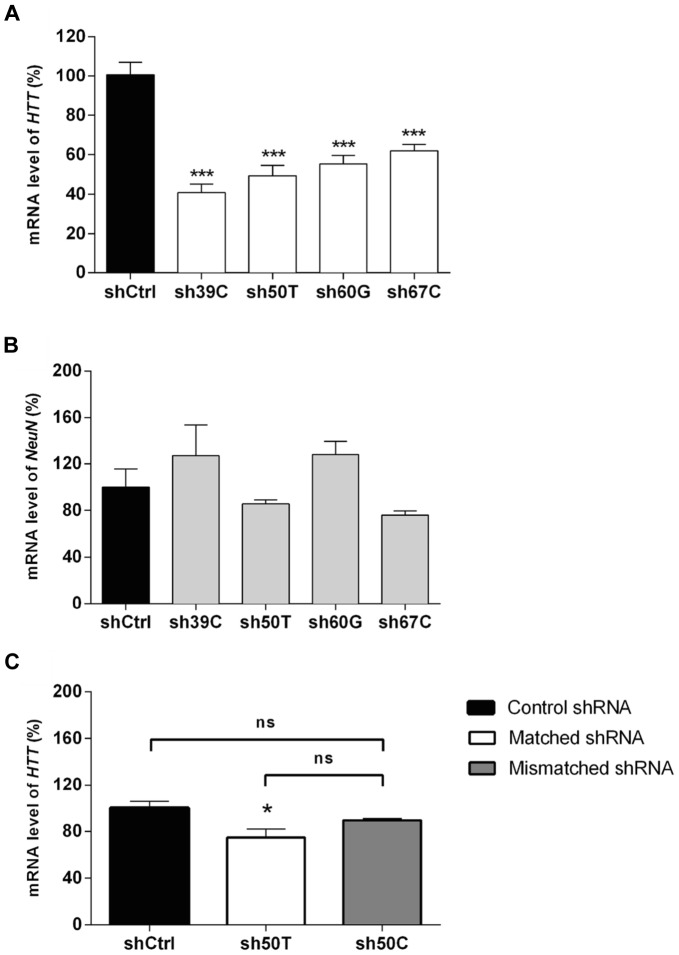
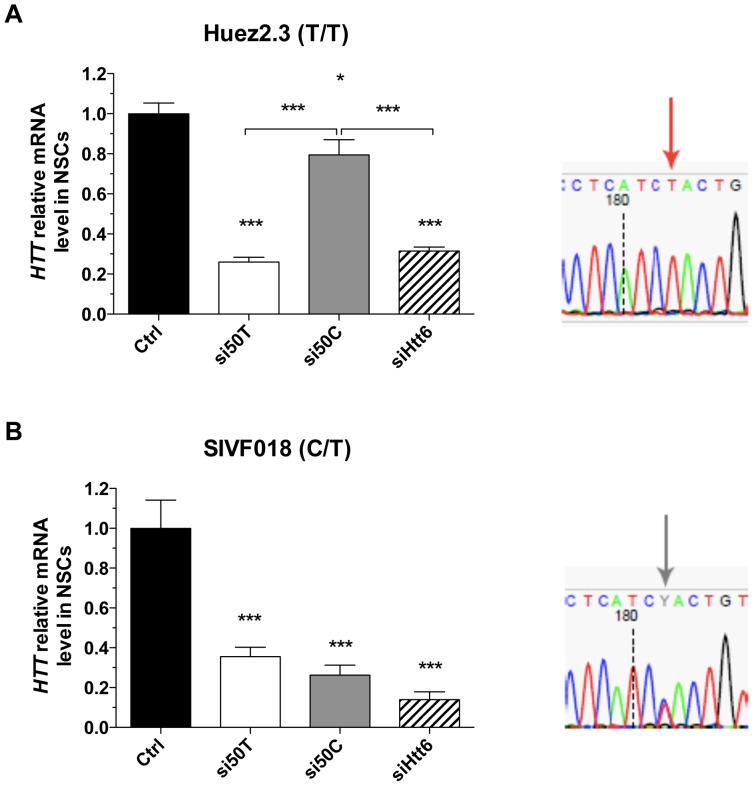
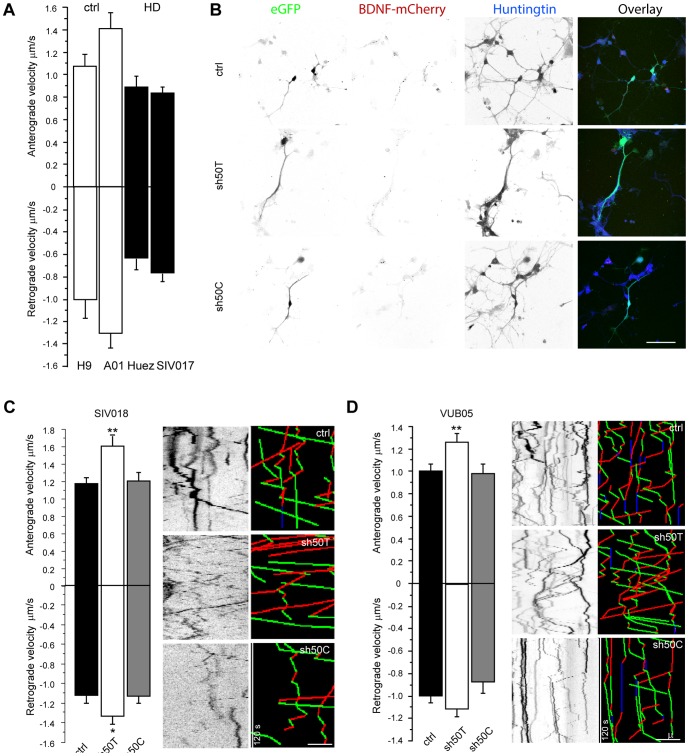
Huntington's disease (HD) is an autosomal dominant neurodegenerative disorder resulting from polyglutamine expansion in the huntingtin (HTT) protein and for which there is no cure. Although suppression of both wild type and mutant HTT expression by RNA interference is a promising therapeutic strategy, a selective silencing of mutant HTT represents the safest approach preserving WT HTT expression and functions. We developed small hairpin RNAs (shRNAs) targeting single nucleotide polymorphisms (SNP) present in the HTT gene to selectively target the disease HTT isoform. Most of these shRNAs silenced, efficiently and selectively, mutant HTT in vitro. Lentiviral-mediated infection with the shRNAs led to selective degradation of mutant HTT mRNA and prevented the apparition of neuropathology in HD rat's striatum expressing mutant HTT containing the various SNPs. In transgenic BACHD mice, the mutant HTT allele was also silenced by this approach, further demonstrating the potential for allele-specific silencing. Finally, the allele-specific silencing of mutant HTT in human embryonic stem cells was accompanied by functional recovery of the vesicular transport of BDNF along microtubules. These findings provide evidence of the therapeutic potential of allele-specific RNA interference for HD.
Conflict of interest statement
Figures
References
-
- Vonsattel JP, Myers RH, Stevens TJ, Ferrante RJ, Bird ED, et al. (1985) Neuropathological classification of Huntington's disease. J Neuropathol Exp Neurol 44: 559–577. - PubMed
-
- The, Huntington's disease collaborative research (1993) A novel gene containing a trinucleotide repeat that is expanded and unstable on Huntington's disease chromosome. Cell 72: 971–983. - PubMed
-
- DiFiglia M, Sapp E, Chase KO, Davies SW, Bates GP, et al. (1997) Aggregation of huntingtin in neuronal intranuclear inclusions and dystrophic neurites in brain. Science 277: 1990–1993. - PubMed
-
- Zhang Y, Friedlander RM (2011) Using non-coding small RNAs to develop therapies for Huntington's disease. Gene Ther 18: 1139–1149. - PubMed
-
- Drouet V, Perrin V, Hassig R, Dufour N, Auregan G, et al. (2009) Sustained effects of nonallele-specific Huntingtin silencing. Ann Neurol 65: 276–285. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
