

Cofactor-dependent conformational heterogeneity of GAD65 and its role in autoimmunity and neurotransmitter homeostasis
- PMID: 24927554
- PMCID: PMC4078817
- DOI: 10.1073/pnas.1403182111
Cofactor-dependent conformational heterogeneity of GAD65 and its role in autoimmunity and neurotransmitter homeostasis
Abstract
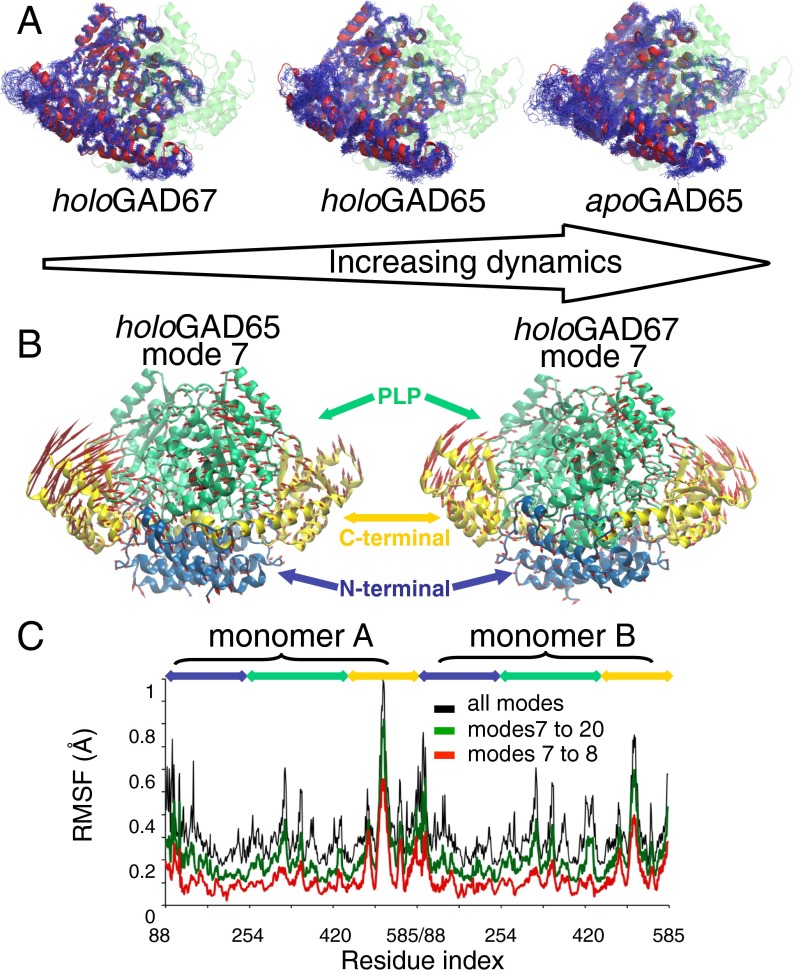
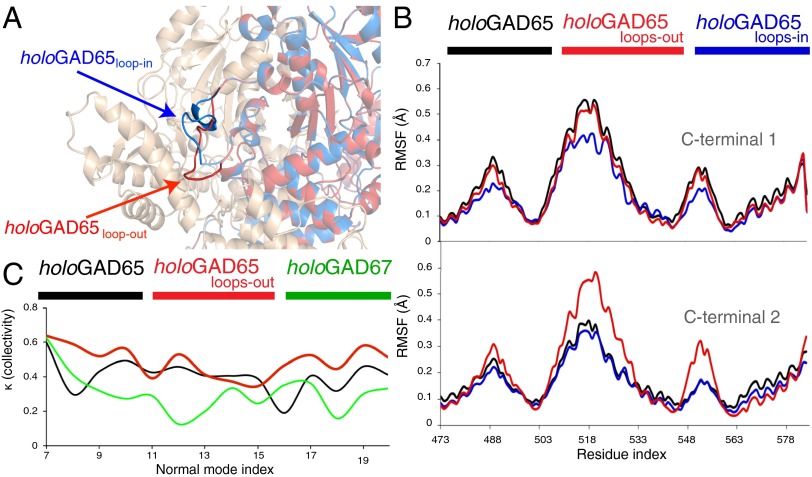
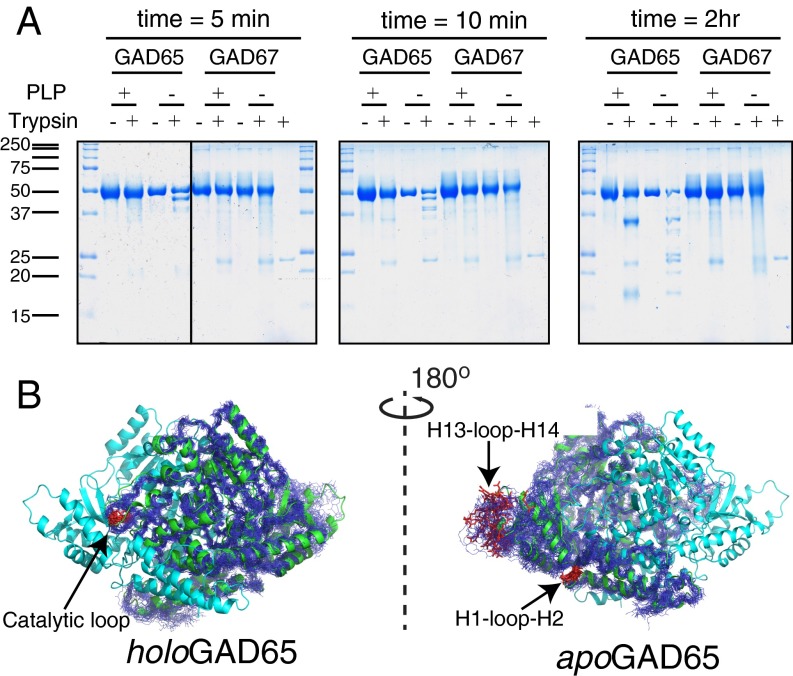
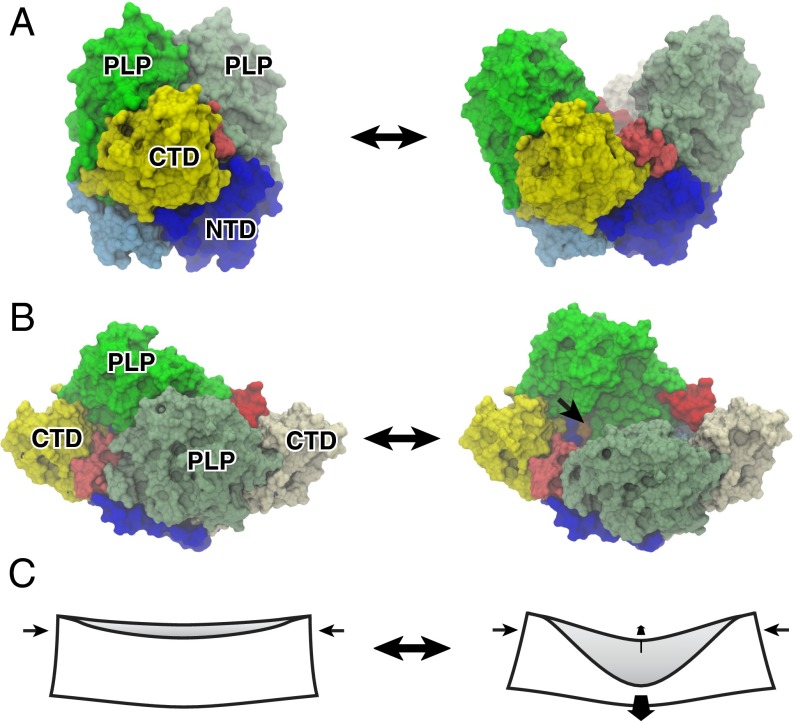
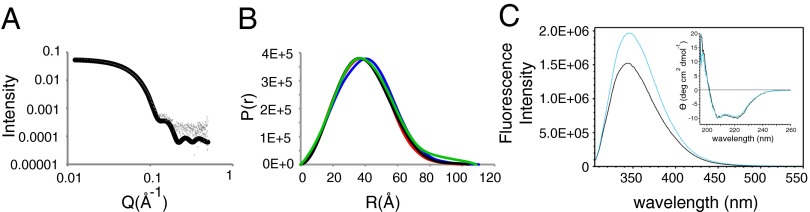
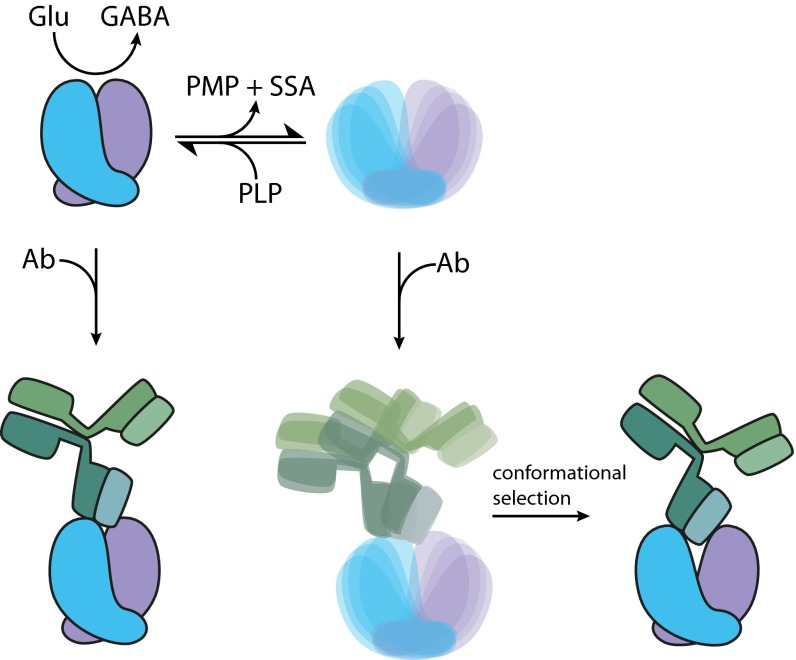
The human neuroendocrine enzyme glutamate decarboxylase (GAD) catalyses the synthesis of the inhibitory neurotransmitter gamma-aminobutyric acid (GABA) using pyridoxal 5'-phosphate as a cofactor. GAD exists as two isoforms named according to their respective molecular weights: GAD65 and GAD67. Although cytosolic GAD67 is typically saturated with the cofactor (holoGAD67) and constitutively active to produce basal levels of GABA, the membrane-associated GAD65 exists mainly as the inactive apo form. GAD65, but not GAD67, is a prevalent autoantigen, with autoantibodies to GAD65 being detected at high frequency in patients with autoimmune (type 1) diabetes and certain other autoimmune disorders. The significance of GAD65 autoinactivation into the apo form for regulation of neurotransmitter levels and autoantibody reactivity is not understood. We have used computational and experimental approaches to decipher the nature of the holo → apo conversion in GAD65 and thus, its mechanism of autoinactivation. Molecular dynamics simulations of GAD65 reveal coupling between the C-terminal domain, catalytic loop, and pyridoxal 5'-phosphate-binding domain that drives structural rearrangement, dimer opening, and autoinactivation, consistent with limited proteolysis fragmentation patterns. Together with small-angle X-ray scattering and fluorescence spectroscopy data, our findings are consistent with apoGAD65 existing as an ensemble of conformations. Antibody-binding kinetics suggest a mechanism of mutually induced conformational changes, implicating the flexibility of apoGAD65 in its autoantigenicity. Although conformational diversity may provide a mechanism for cofactor-controlled regulation of neurotransmitter biosynthesis, it may also come at a cost of insufficient development of immune self-tolerance that favors the production of GAD65 autoantibodies.
Keywords: GABA biosynthesis; autoepitopes; conformational dynamics; immunogenicity; normal mode analysis.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Erlander MG, Tillakaratne NJ, Feldblum S, Patel N, Tobin AJ. Two genes encode distinct glutamate decarboxylases. Neuron. 1991;7(1):91–100. - PubMed
-
- Sandmeier E, Hale TI, Christen P. Multiple evolutionary origin of pyridoxal-5'-phosphate-dependent amino acid decarboxylases. Eur J Biochem. 1994;221(3):997–1002. - PubMed
-
- Martin DL, Rimvall K. Regulation of gamma-aminobutyric acid synthesis in the brain. J Neurochem. 1993;60(2):395–407. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
