

Identification of Mn(II)-oxidizing bacteria from a low-pH contaminated former uranium mine
- PMID: 24928873
- PMCID: PMC4135758
- DOI: 10.1128/AEM.01296-14
Identification of Mn(II)-oxidizing bacteria from a low-pH contaminated former uranium mine
Abstract
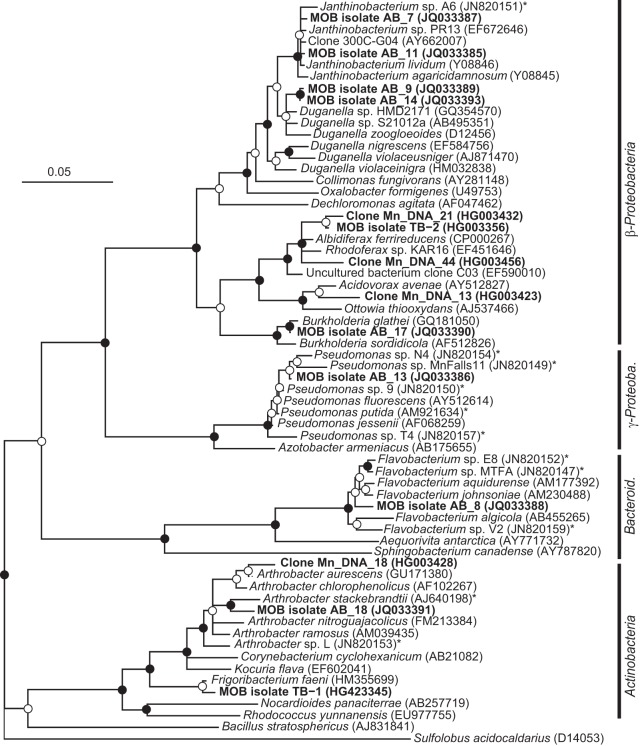
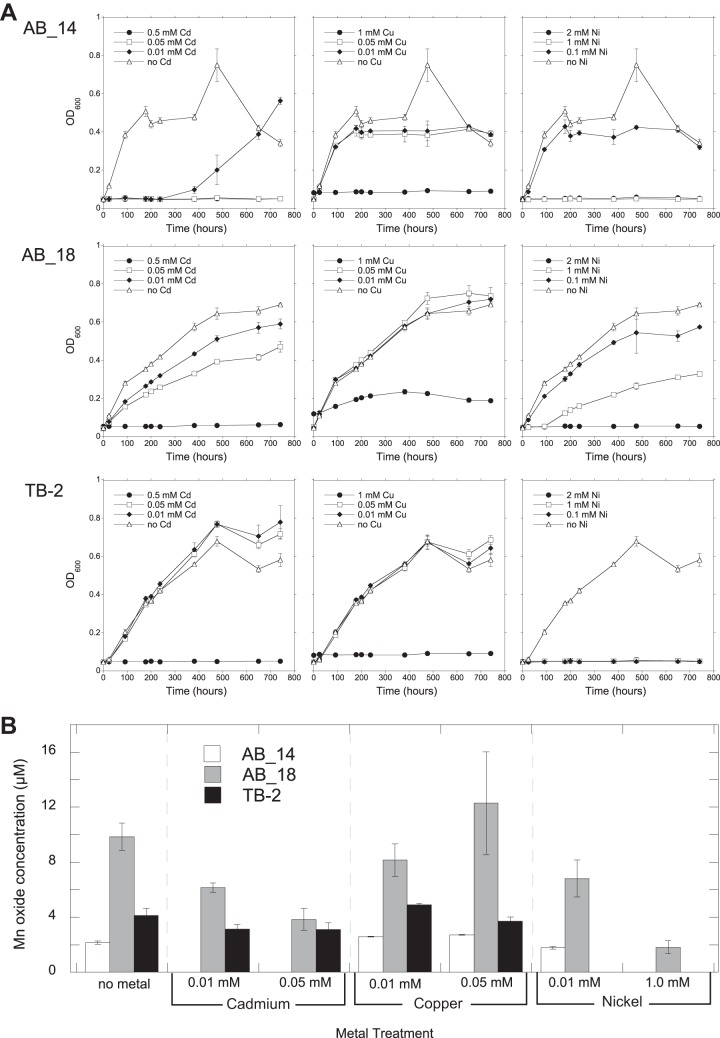
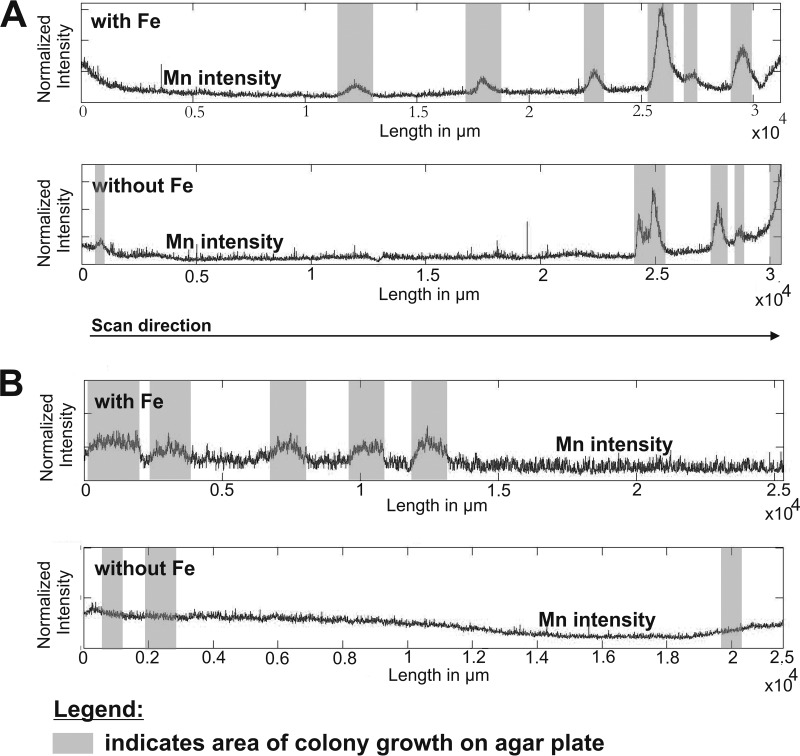
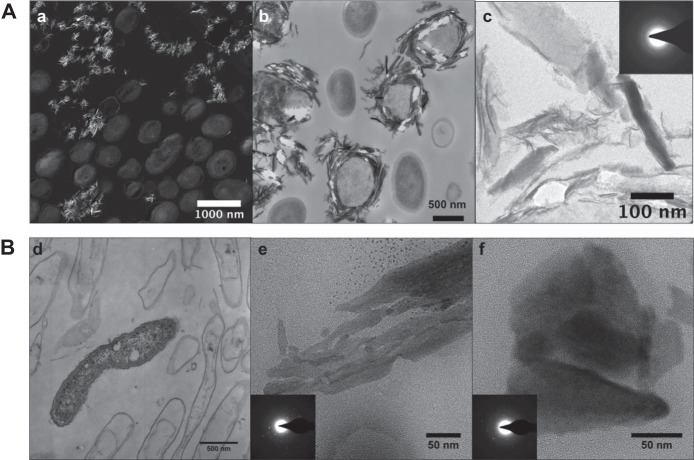
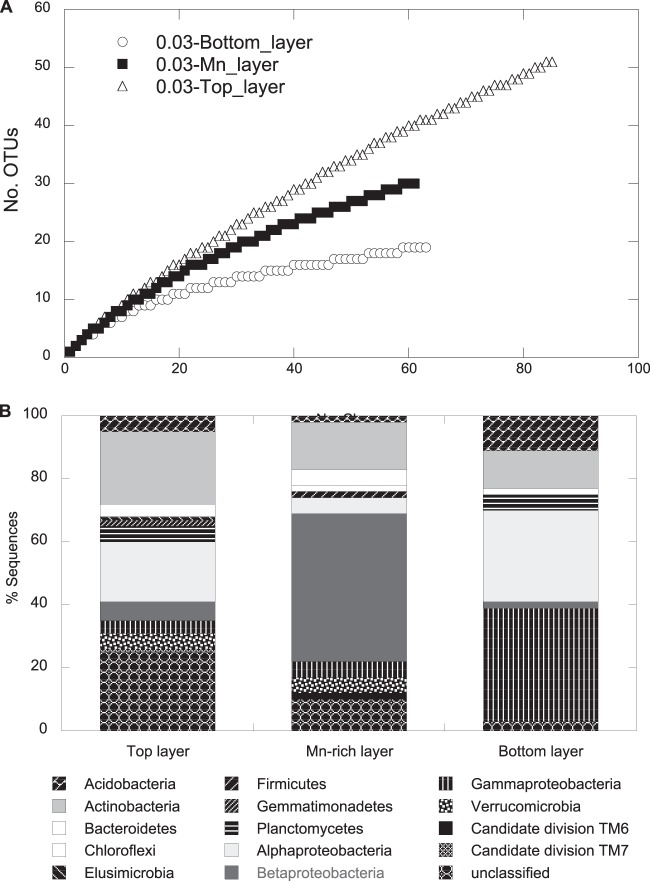
Biological Mn oxidation is responsible for producing highly reactive and abundant Mn oxide phases in the environment that can mitigate metal contamination. However, little is known about Mn oxidation in low-pH environments, where metal contamination often is a problem as the result of mining activities. We isolated two Mn(II)-oxidizing bacteria (MOB) at pH 5.5 (Duganella isolate AB_14 and Albidiferax isolate TB-2) and nine strains at pH 7 from a former uranium mining site. Isolate TB-2 may contribute to Mn oxidation in the acidic Mn-rich subsoil, as a closely related clone represented 16% of the total community. All isolates oxidized Mn over a small pH range, and isolates from low-pH samples only oxidized Mn below pH 6. Two strains with different pH optima differed in their Fe requirements for Mn oxidation, suggesting that Mn oxidation by the strain found at neutral pH was linked to Fe oxidation. Isolates tolerated Ni, Cu, and Cd and produced Mn oxides with similarities to todorokite and birnessite, with the latter being present in subsurface layers where metal enrichment was associated with Mn oxides. This demonstrates that MOB can be involved in the formation of biogenic Mn oxides in both moderately acidic and neutral pH environments.
Copyright © 2014, American Society for Microbiology. All Rights Reserved.
Figures
References
-
- Morgan JJ. 2005. Kinetics of reaction between O2 and Mn(II) species in aqueous solutions. Geochim. Cosmochim. Acta 69:35–48. 10.1016/j.gca.2004.06.013 - DOI
-
- Tebo BM, Bargar JR, Clement BG, Dick GJ, Murray KJ, Parker D, Verity R, Webb SM. 2004. Biogenic manganese oxides: properties and mechanisms of formation. Annu. Rev. Earth Planet. Sci. 32:287–328. 10.1146/annurev.earth.32.101802.120213 - DOI
-
- Tebo B, Clement B, Dick G. 2007. Biotransformations of manganese, p 1223–1238 In Hurst CJ. (ed), Manual of environmental microbiology, 3rd ed. ASM Press, Washington, DC
-
- Nealson KH. 2006. The manganese-oxidizing bacteria, p 222–231 In Balows A, Trüper HG, Dworkin M, Harder W, Schleifer K-H. (ed), The prokaryotes. Springer, New York, NY
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
