

Real-time detection of CTL function reveals distinct patterns of caspase activation mediated by Fas versus granzyme B
- PMID: 24928990
- PMCID: PMC4107314
- DOI: 10.4049/jimmunol.1301668
Real-time detection of CTL function reveals distinct patterns of caspase activation mediated by Fas versus granzyme B
Abstract
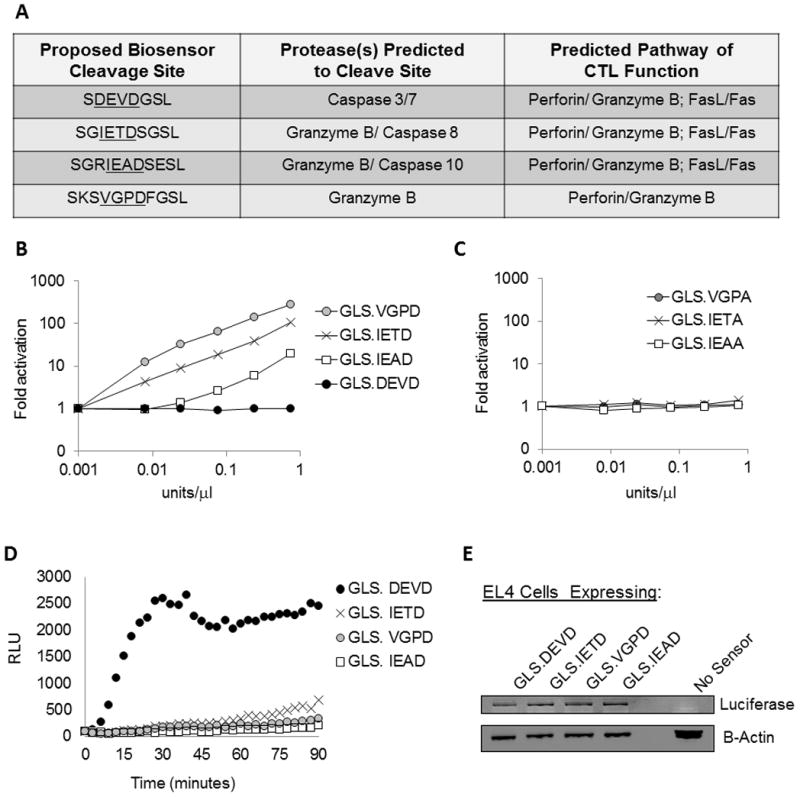
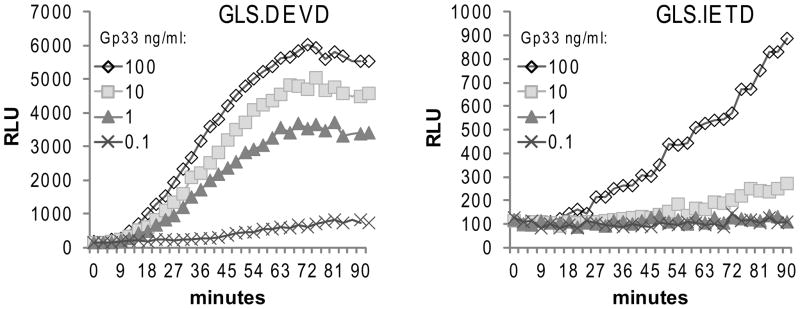
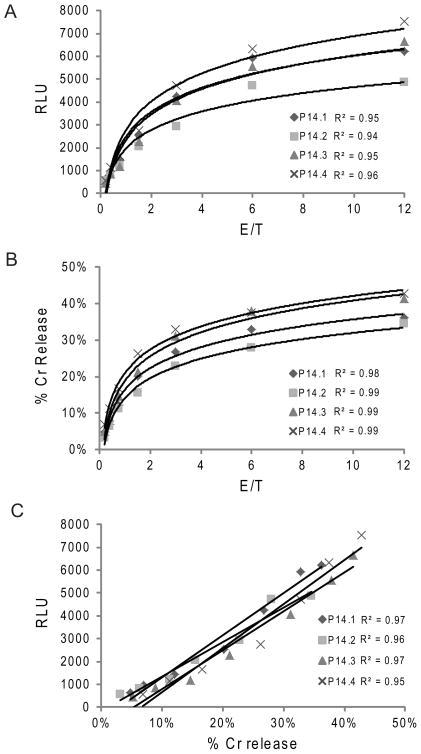
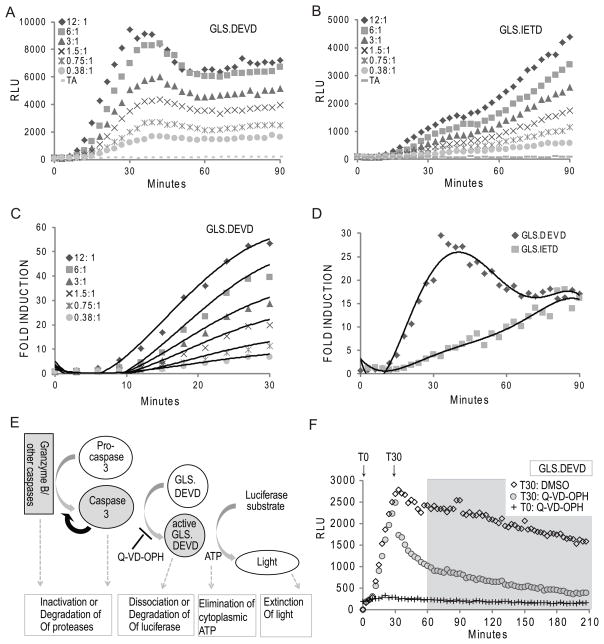
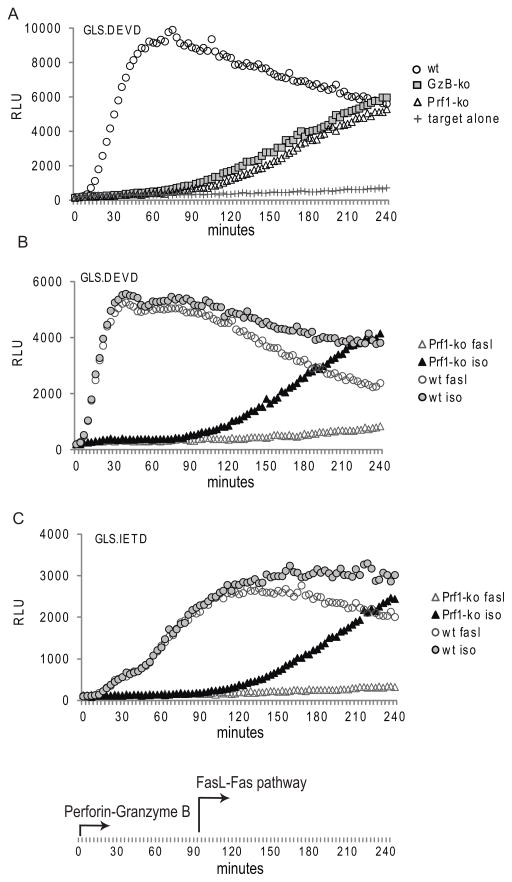
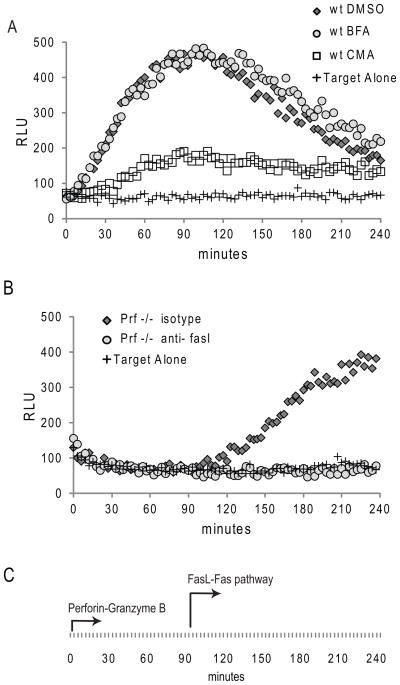
Activation of caspase-mediated apoptosis is reported to be a hallmark of both granzyme B- and Fas-mediated pathways of killing by CTLs; however, the kinetics of caspase activation remain undefined owing to an inability to monitor target cell-specific apoptosis in real time. We have overcome this limitation by developing a novel biosensor assay that detects continuous, protease-specific activity in target cells. Biosensors were engineered from a circularly permuted luciferase, linked internally by either caspase 3/7 or granzyme B/caspase 8 cleavage sites, thus allowing activation upon proteolytic cleavage by the respective proteases. Coincubation of murine CTLs with target cells expressing either type of biosensor led to a robust luminescent signal within minutes of cell contact. The signal was modulated by the strength of TCR signaling, the ratio of CTL/target cells, and the type of biosensor used. Additionally, the luciferase signal at 30 min correlated with target cell death, as measured by a (51)Cr-release assay. The rate of caspase 3/7 biosensor activation was unexpectedly rapid following granzyme B- compared with Fas-mediated signal induction in murine CTLs; the latter appeared gradually after a 90-min delay in perforin- or granzyme B-deficient CTLs. Remarkably, the Fas-dependent, caspase 3/7 biosensor signal induced by perforin-deficient human CTLs was also detectable after a 90-min delay when measured by redirected killing. Thus, we have used a novel, real-time assay to demonstrate the distinct pattern of caspase activation induced by granzyme B versus Fas in human and murine CTLs.
Copyright © 2014 by The American Association of Immunologists, Inc.
Figures
References
-
- de Saint Basile G, Menasche G, Fischer A. Molecular mechanisms of biogenesis and exocytosis of cytotoxic granules. Nat Rev Immunol. 2010;10:568–579. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
