

Dicer-microRNA-Myc circuit promotes transcription of hundreds of long noncoding RNAs
- PMID: 24929436
- PMCID: PMC5509563
- DOI: 10.1038/nsmb.2842
Dicer-microRNA-Myc circuit promotes transcription of hundreds of long noncoding RNAs
Abstract
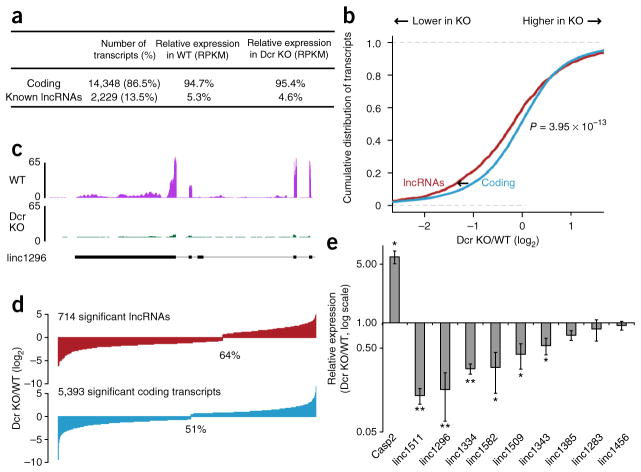
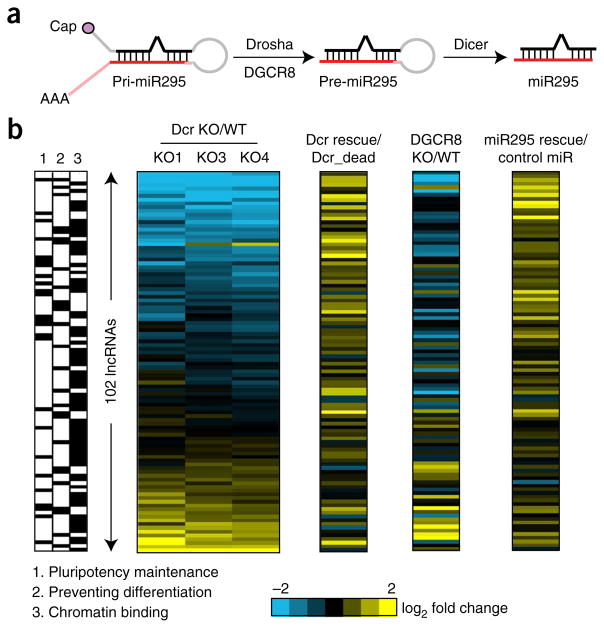
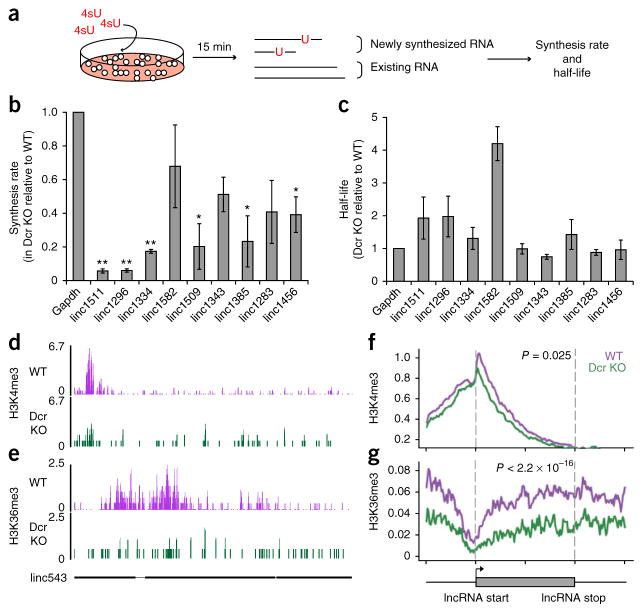
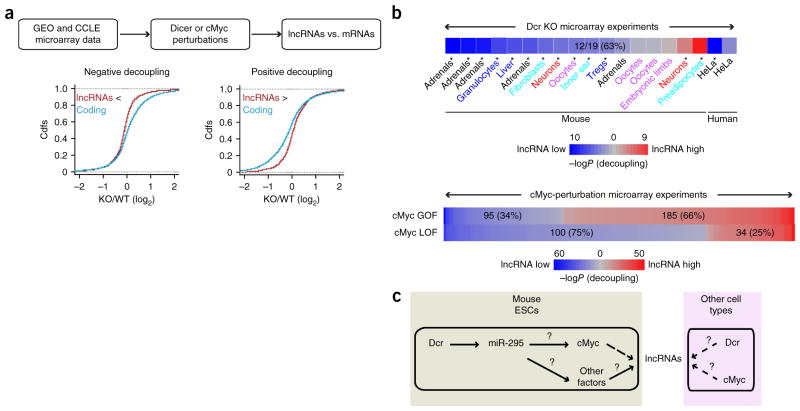
Long noncoding RNAs (lncRNAs) are important regulators of cell fate, yet little is known about mechanisms controlling lncRNA expression. Here we show that transcription is quantitatively different for lncRNAs and mRNAs--as revealed by deficiency of Dicer (Dcr), a key RNase that generates microRNAs (miRNAs). Dcr loss in mouse embryonic stem cells led unexpectedly to decreased levels of hundreds of lncRNAs. The canonical Dgcr8-Dcr-miRNA pathway is required for robust lncRNA transcriptional initiation and elongation. Computational and genetic epistasis analyses demonstrated that Dcr activation of the oncogenic transcription factor cMyc is partly responsible for lncRNA expression. A quantitative metric of mRNA-lncRNA decoupling revealed that Dcr and cMyc differentially regulate lncRNAs versus mRNAs in diverse cell types and in vivo. Thus, numerous lncRNAs may be modulated as a class in development and disease, notably where Dcr and cMyc act.
Conflict of interest statement
The authors declare no competing financial interests.
Figures

References
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
