

The parasitophorous vacuole membrane of Toxoplasma gondii is targeted for disruption by ubiquitin-like conjugation systems of autophagy
- PMID: 24931121
- PMCID: PMC4107903
- DOI: 10.1016/j.immuni.2014.05.006
The parasitophorous vacuole membrane of Toxoplasma gondii is targeted for disruption by ubiquitin-like conjugation systems of autophagy
Abstract
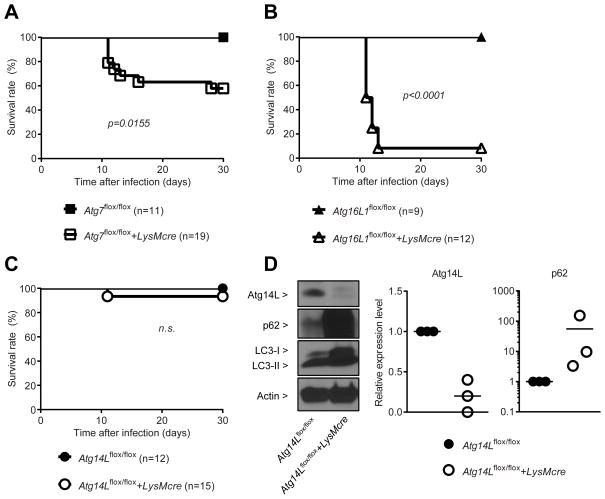
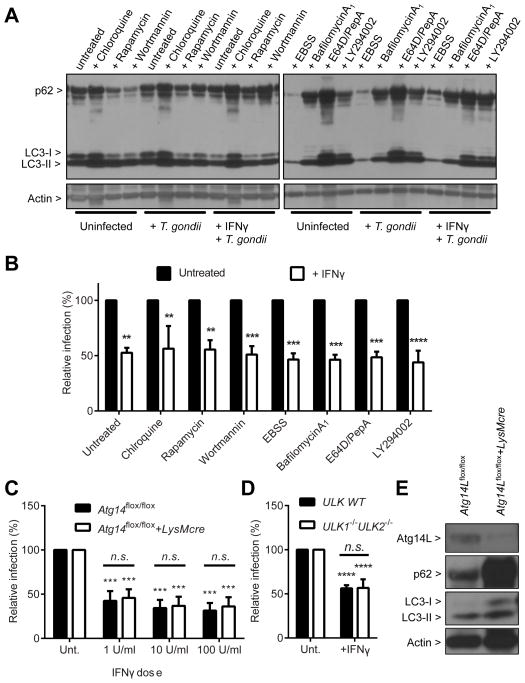
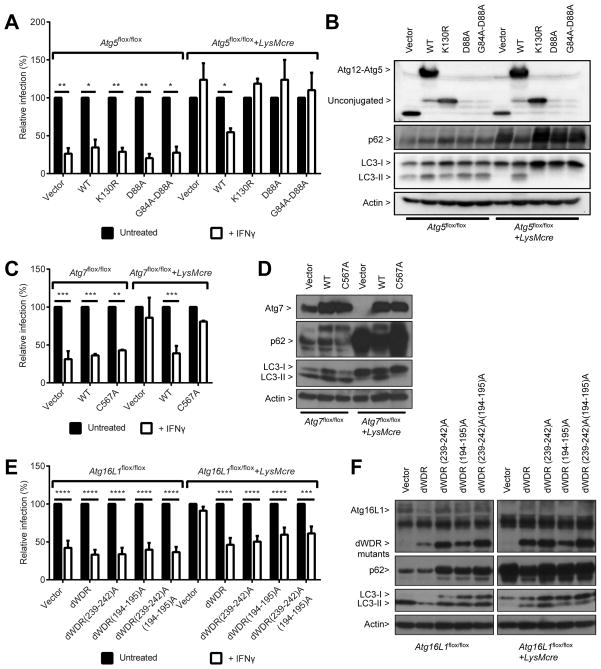
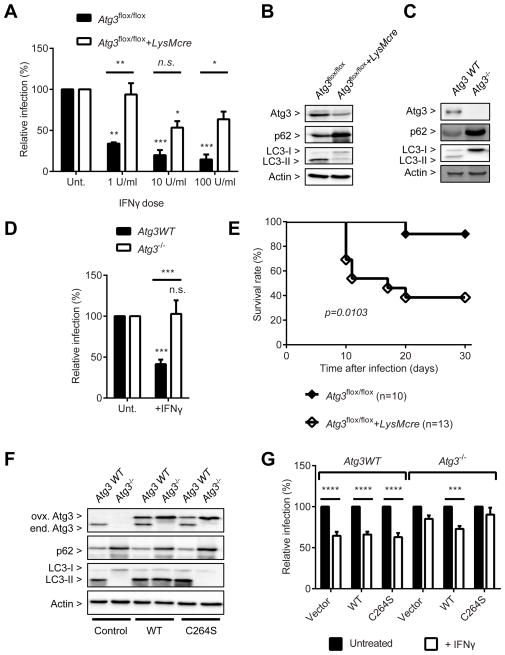
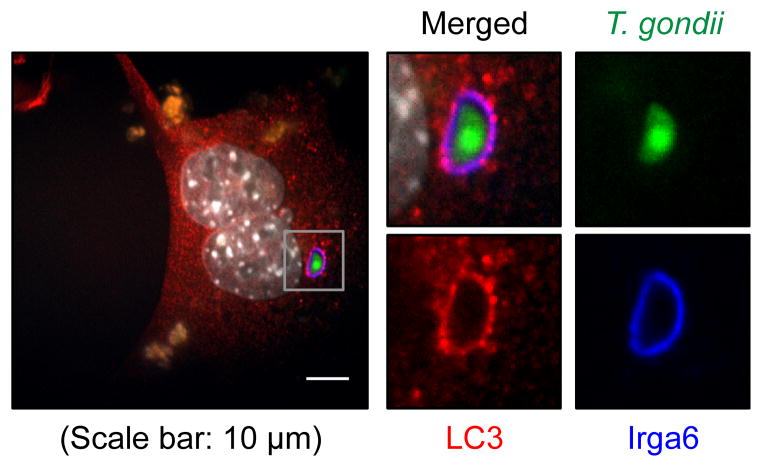
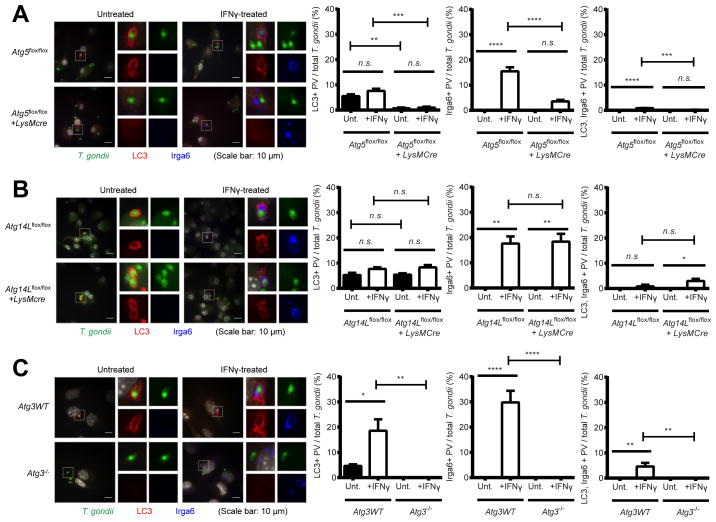
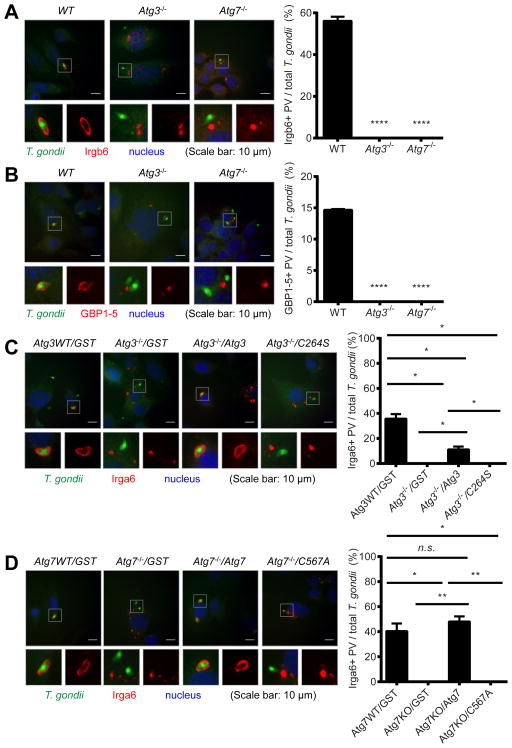
Autophagy is a lysosomal degradation pathway that is important in cellular homeostasis. Prior work showed a key role for the autophagy related 5 (Atg5) in resistance to Toxoplasma gondii. Here we show that the cassette of autophagy proteins involved in the conjugation of microtubule-associated protein 1 light chain 3 (LC3) to phosphatidylethanolamine, including Atg7, Atg3, and the Atg12-Atg5-Atg16L1 complex play crucial roles in the control of T. gondii in vitro and in vivo. In contrast, pharmacologic modulation of the degradative autophagy pathway or genetic deletion of other essential autophagy genes had no substantial effects. Rather the conjugation system was required for targeting of LC3 and interferon-γ effectors onto the vacuolar membrane of T. gondii and its consequent disruption. These data suggest that the ubiquitin-like conjugation systems that reorganize intracellular membranes during canonical autophagy are necessary for proper targeting of immune effectors to the intracellular vacuole membranes utilized by pathogens.
Copyright © 2014 Elsevier Inc. All rights reserved.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
