

Multicilin drives centriole biogenesis via E2f proteins
- PMID: 24934224
- PMCID: PMC4083089
- DOI: 10.1101/gad.243832.114
Multicilin drives centriole biogenesis via E2f proteins
Abstract
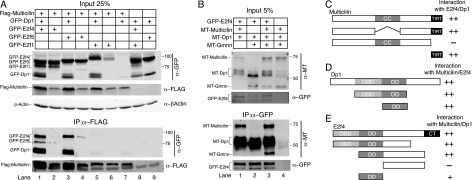
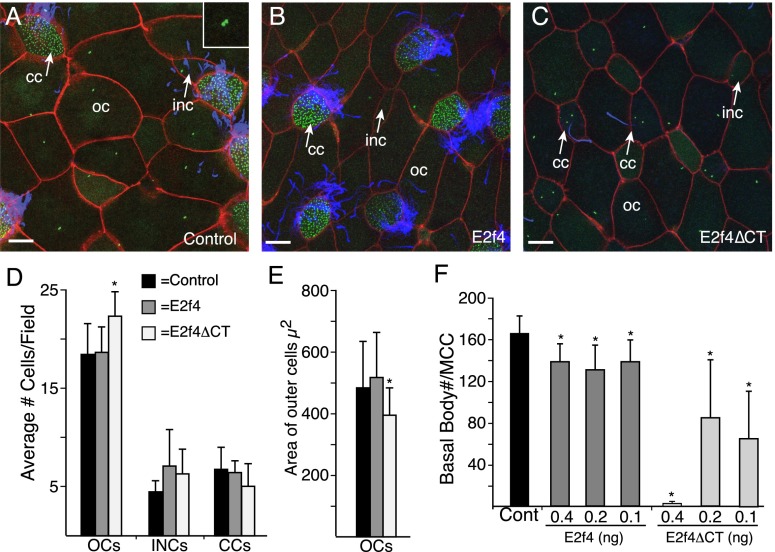
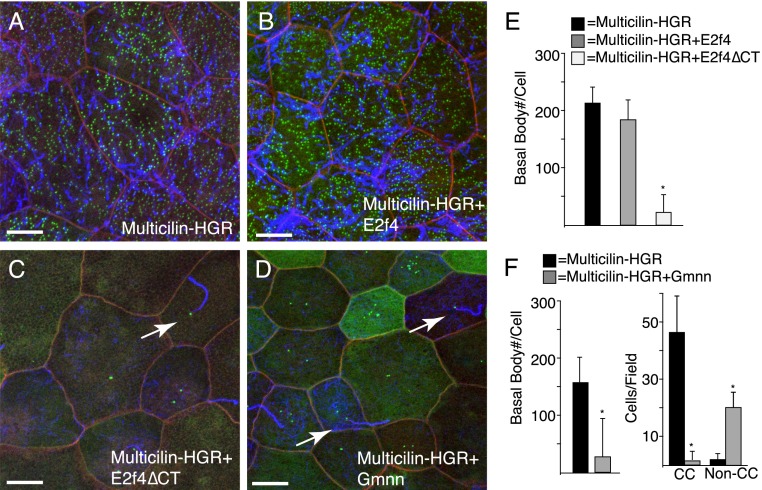
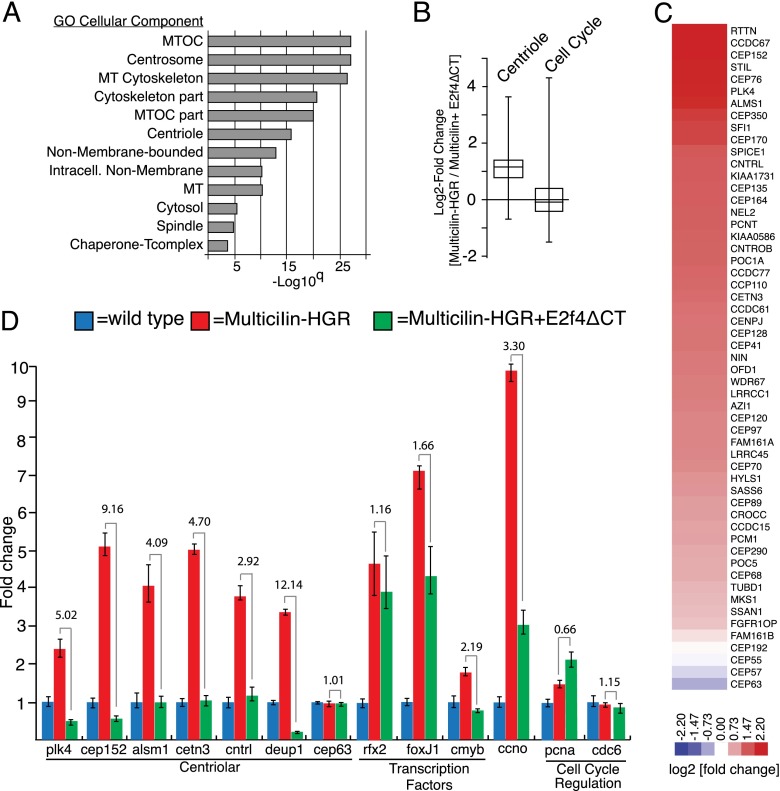
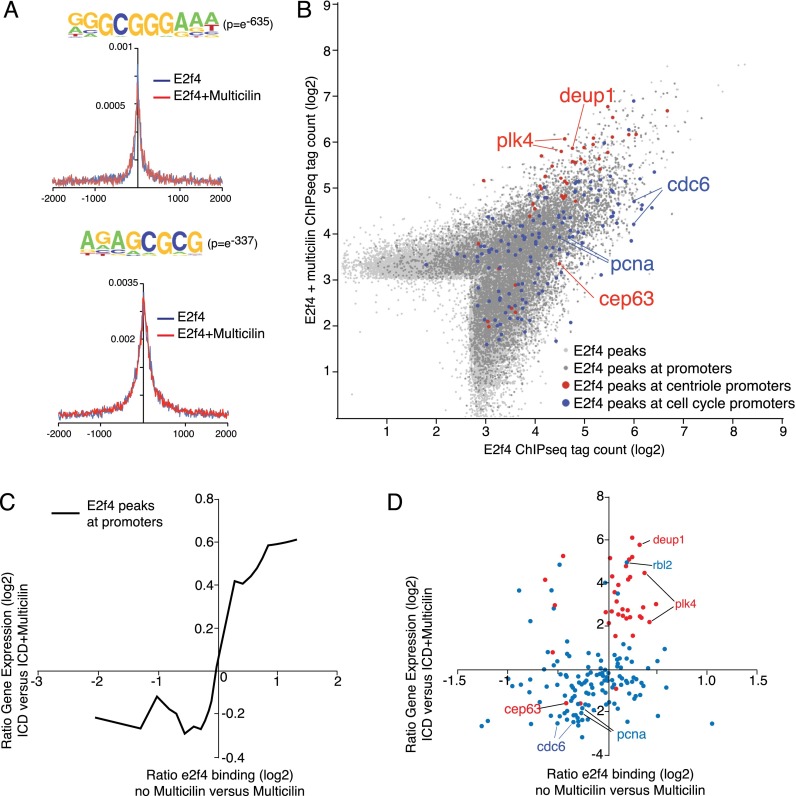
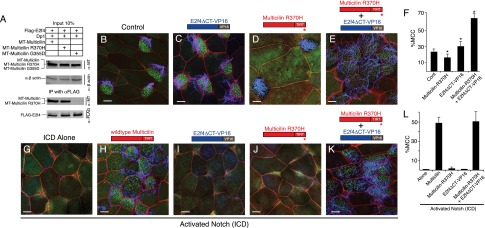
Multiciliate cells employ hundreds of motile cilia to produce fluid flow, which they nucleate and extend by first assembling hundreds of centrioles. In most cells, entry into the cell cycle allows centrioles to undergo a single round of duplication, but in differentiating multiciliate cells, massive centriole assembly occurs in G0 by a process initiated by a small coiled-coil protein, Multicilin. Here we show that Multicilin acts by forming a ternary complex with E2f4 or E2f5 and Dp1 that binds and activates most of the genes required for centriole biogenesis, while other cell cycle genes remain off. This complex also promotes the deuterosome pathway of centriole biogenesis by activating the expression of deup1 but not its paralog, cep63. Finally, we show that this complex is disabled by mutations in human Multicilin that cause a severe congenital mucociliary clearance disorder due to reduced generation of multiple cilia. By coopting the E2f regulation of cell cycle genes, Multicilin drives massive centriole assembly in epithelial progenitors in a manner required for multiciliate cell differentiation.
Keywords: centrioles; e2f4; multiciliate cells.
© 2014 Ma et al.; Published by Cold Spring Harbor Laboratory Press.
Figures
Comment in
-
Multiciliogenesis: multicilin directs transcriptional activation of centriole formation.Curr Biol. 2014 Aug 18;24(16):R746-9. doi: 10.1016/j.cub.2014.07.006. Curr Biol. 2014. PMID: 25137586
References
-
- Bettencourt-Dias M, Rodrigues-Martins A, Carpenter L, Riparbelli M, Lehmann L, Gatt MK, Carmo N, Balloux F, Callaini G, Glover DM 2005. SAK/PLK4 is required for centriole duplication and flagella development. Curr Biol 15: 2199–2207 - PubMed
-
- Boon M, Wallmeier J, Ma L, Loges NT, Jaspers M, Olbrich H, Dougherty G, Werner C, Amirav I, Hevroni A, et al. 2014. Mutations in MCIDAS result in a congenital mucociliary clearance disorder with reduced generation of multiple motile cilia (RGMC). Nat Commun (in press) - PubMed
-
- Carroll SB 2008. Evo-devo and an expanding evolutionary synthesis: a genetic theory of morphological evolution. Cell 134: 25–36 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases