

Differences in nucleation behavior underlie the contrasting aggregation kinetics of the Aβ40 and Aβ42 peptides
- PMID: 24938782
- PMCID: PMC4084462
- DOI: 10.1073/pnas.1401564111
Differences in nucleation behavior underlie the contrasting aggregation kinetics of the Aβ40 and Aβ42 peptides
Abstract
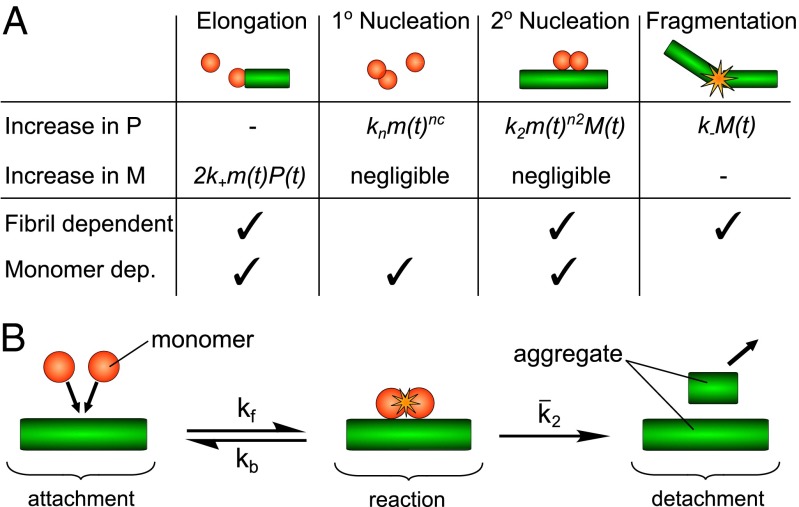
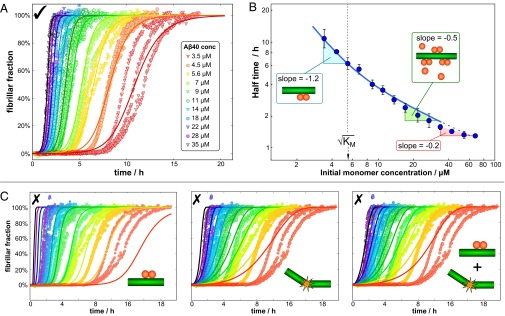
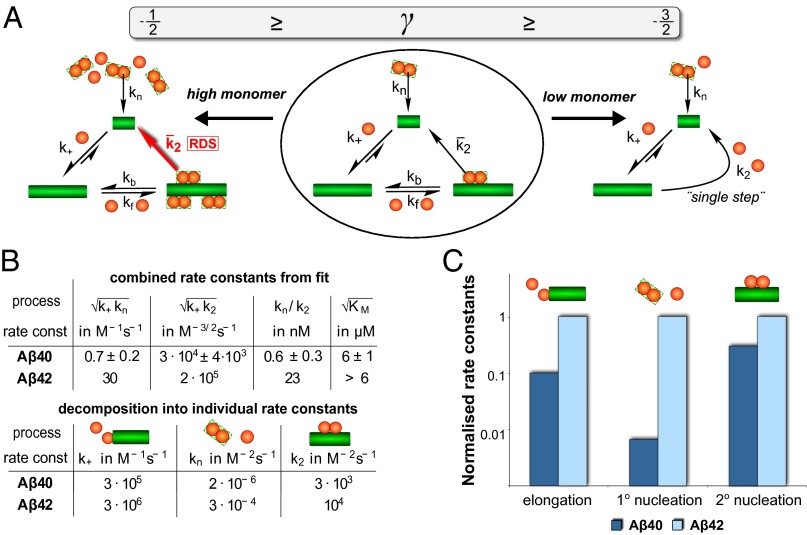
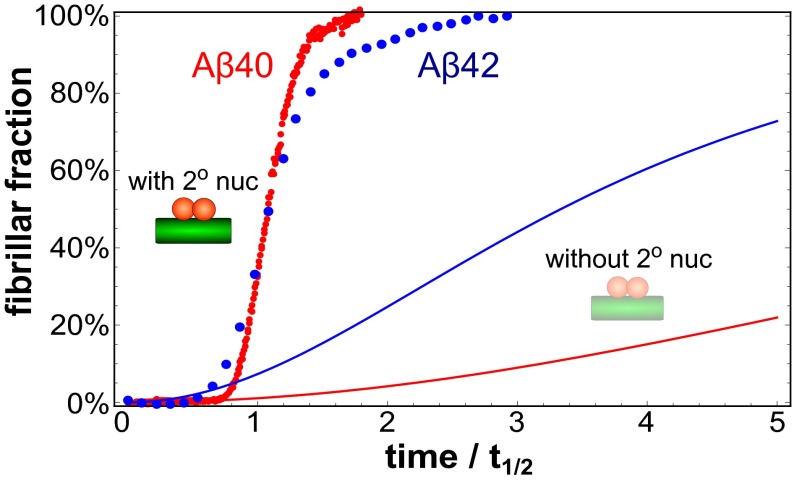
The two major forms of the amyloid-beta (Aβ) peptide found in plaques in patients suffering from Alzheimer's disease, Aβ40 and Aβ42, only differ by two amino acids in the C-terminal region, yet they display markedly different aggregation behavior. The origins of these differences have remained challenging to connect to specific molecular-level processes underlying the aggregation reaction. In this paper we use a general strategy to apply the conventional workflow of chemical kinetics to the aggregation of the Aβ40 peptide to identify the differences between Aβ40 and Aβ42 in terms of the microscopic determinants of the aggregation reaction. Our results reveal that the major source of aggregates in the case of Aβ40 is a fibril-catalyzed nucleation process, the multistep nature of which is evident through its saturation behavior. Moreover, our results show that the significant differences in the observed behavior of the two proteins originate not simply from a uniform increase in all microscopic rates for Aβ42 compared with Aβ40, but rather are due to a shift of more than one order of magnitude in the relative importance of primary nucleation versus fibril-catalyzed secondary nucleation processes. This analysis sheds light on the microscopic determinants of the aggregation behavior of the principal forms of Aβ and outlines a general approach toward achieving an understanding at the molecular level of the aberrant deposition of insoluble peptides in neurodegenerative disorders.
Keywords: aggregation mechanism; neurodegeneration; protein aggregation; rate law.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Dobson CM. Protein folding and misfolding. Nature. 2003;426(6968):884–890. - PubMed
-
- Aguzzi A, Haass C. Games played by rogue proteins in prion disorders and Alzheimer’s disease. Science. 2003;302(5646):814–818. - PubMed
-
- Aguzzi A, O’Connor T. Protein aggregation diseases: Pathogenicity and therapeutic perspectives. Nat Rev Drug Discov. 2010;9(3):237–248. - PubMed
-
- Hardy J, Selkoe DJ. The amyloid hypothesis of Alzheimer’s disease: Progress and problems on the road to therapeutics. Science. 2002;297(5580):353–356. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
